

Transcriptional and epigenetic regulation of T cell hyporesponsiveness
- PMID: 28606939
- PMCID: PMC5557644
- DOI: 10.1189/jlb.2RI0317-097R
Transcriptional and epigenetic regulation of T cell hyporesponsiveness
Abstract
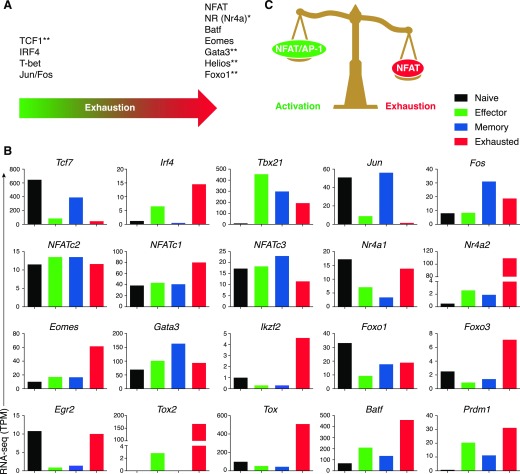
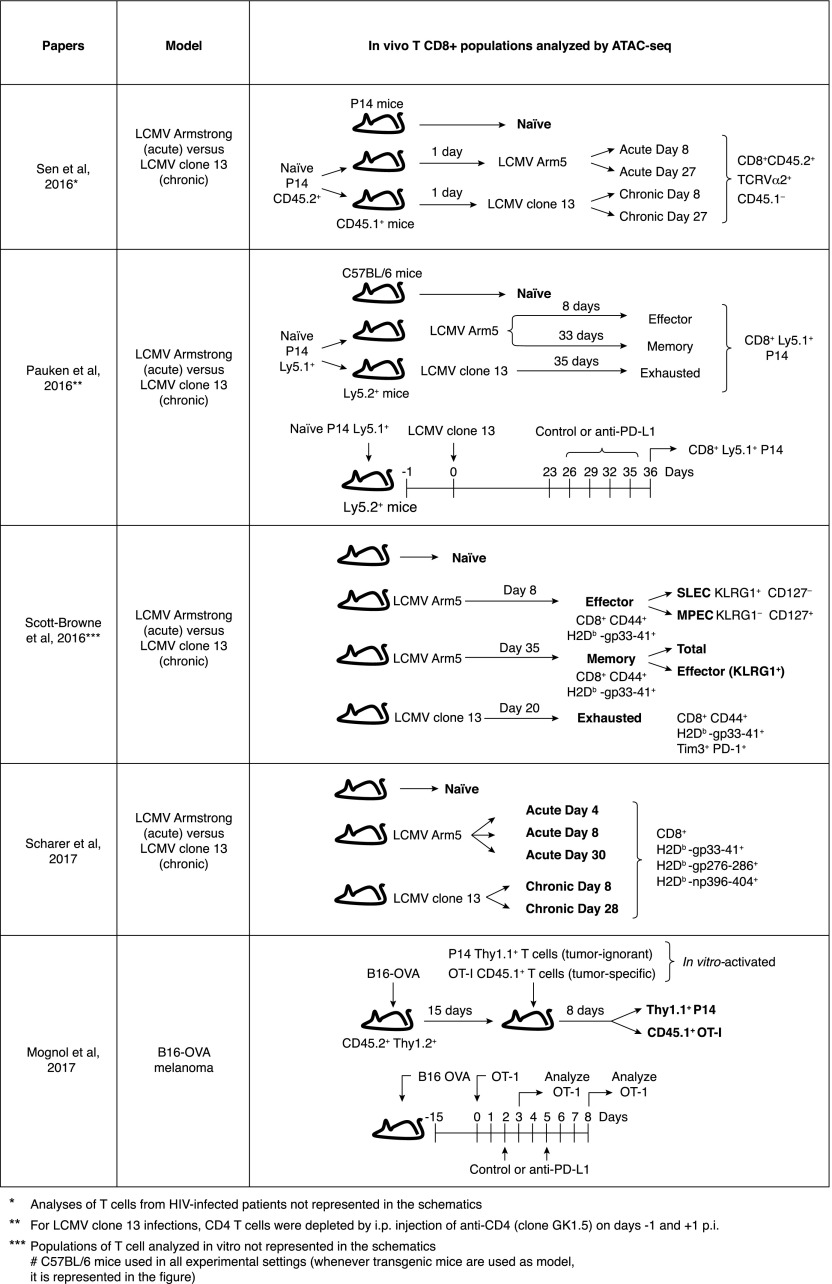
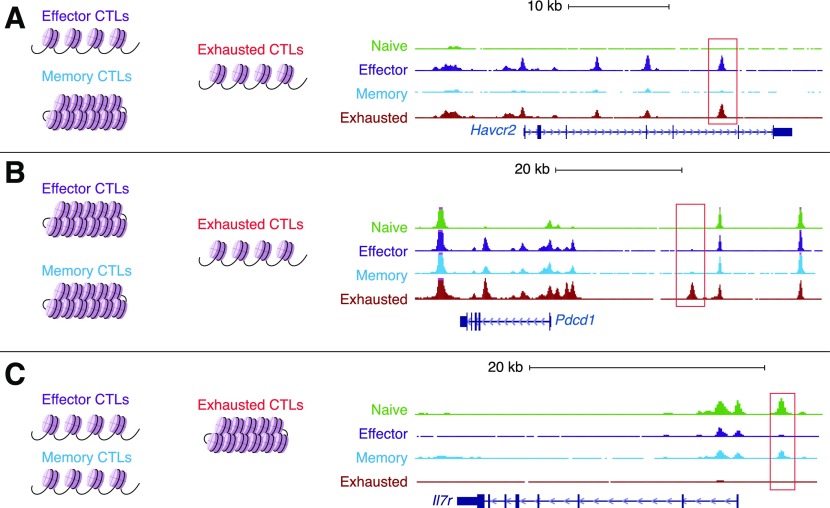
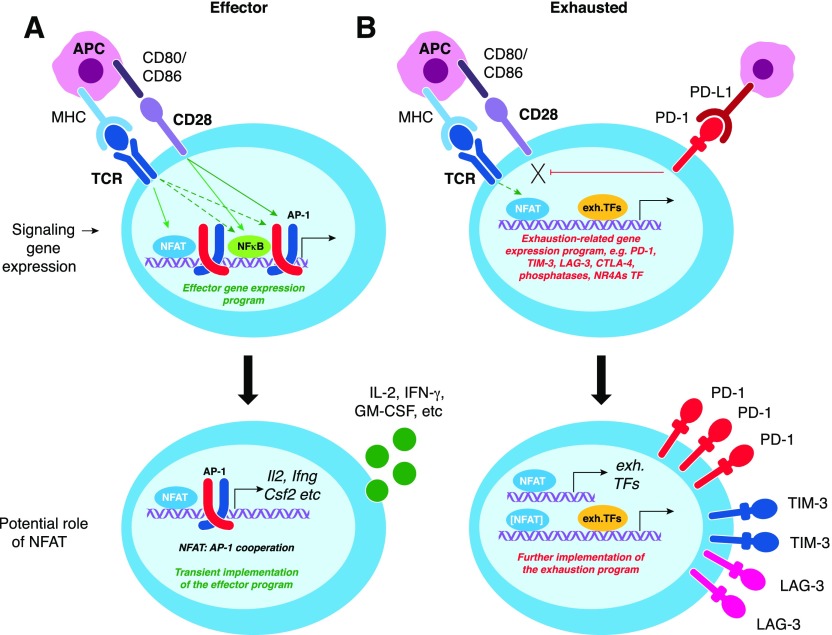
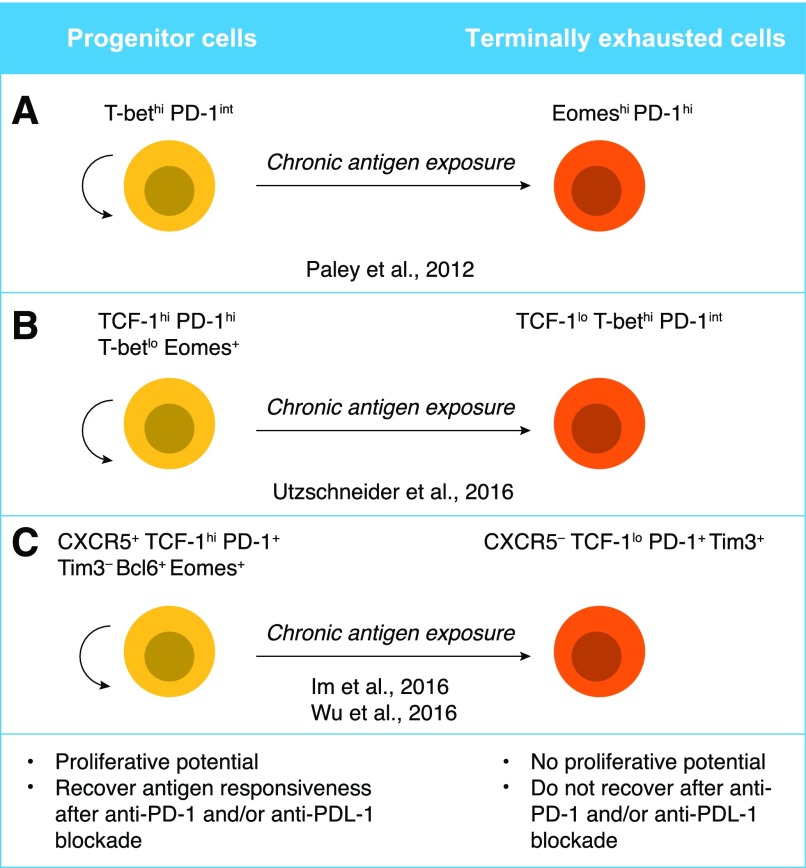
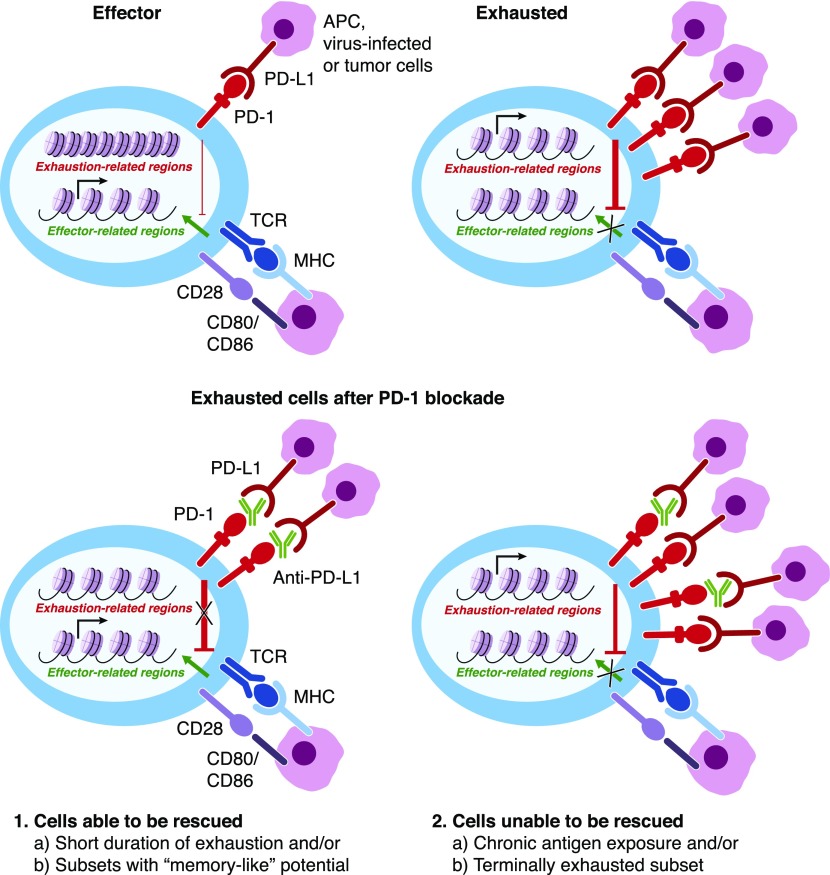
Naive CD8+ T cells differentiate into effector and memory cytolytic T cells (CTLs) during an acute infection. In contrast, in scenarios of persistent antigen stimulation, such as chronic infections and cancer, antigen-specific CTLs show a gradual decrease in effector function, a phenomenon that has been termed CD8+ T cell "exhaustion" or "dysfunction." Another hyporesponsive state, termed "anergy", is observed when T cells are activated in the absence of positive costimulatory signals. Among the many negative regulators induced in hyporesponsive T cells are inhibitory cell-surface receptors, such as PD-1, LAG-3, CTLA-4, and TIM-3; "checkpoint blockade" therapies that involve treatment of patients with cancer with blocking antibodies to those receptors show considerable promise in the clinic because the blocking antibodies can mitigate hyporesponsiveness and promote tumor rejection. In this review, we describe recent advances in our molecular understanding of these hyporesponsive states. We review evidence for the involvement of diverse transcription factors, metabolic programs, and chromatin accessibility changes in hyporesponsive T cells, and we discuss how checkpoint blockade therapies affect the molecular program of CD8+ T cell exhaustion.
Keywords: epigenetic; exhaustion; transcription factors.
© Society for Leukocyte Biology.
Figures
References
-
- Wherry E. J. (2011) T cell exhaustion. Nat. Immunol. 12, 492–499. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
