

The fate and lifespan of human monocyte subsets in steady state and systemic inflammation
- PMID: 28606987
- PMCID: PMC5502436
- DOI: 10.1084/jem.20170355
The fate and lifespan of human monocyte subsets in steady state and systemic inflammation
Abstract
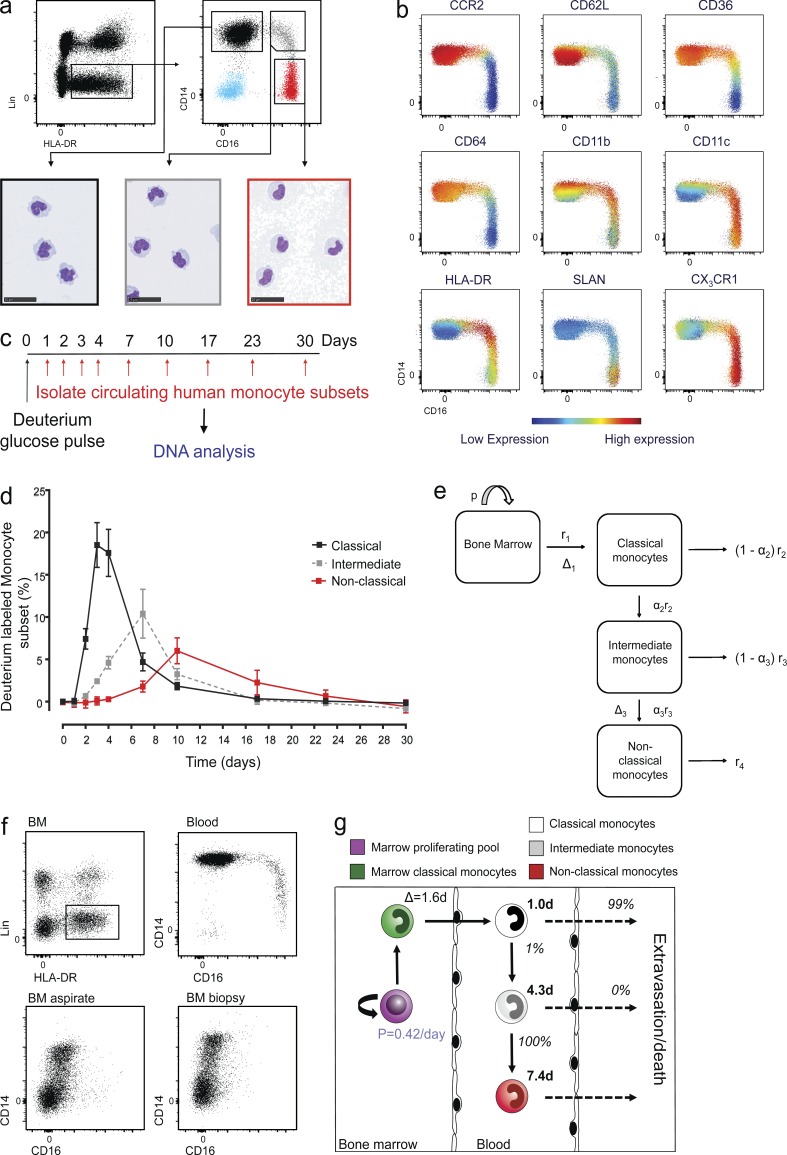
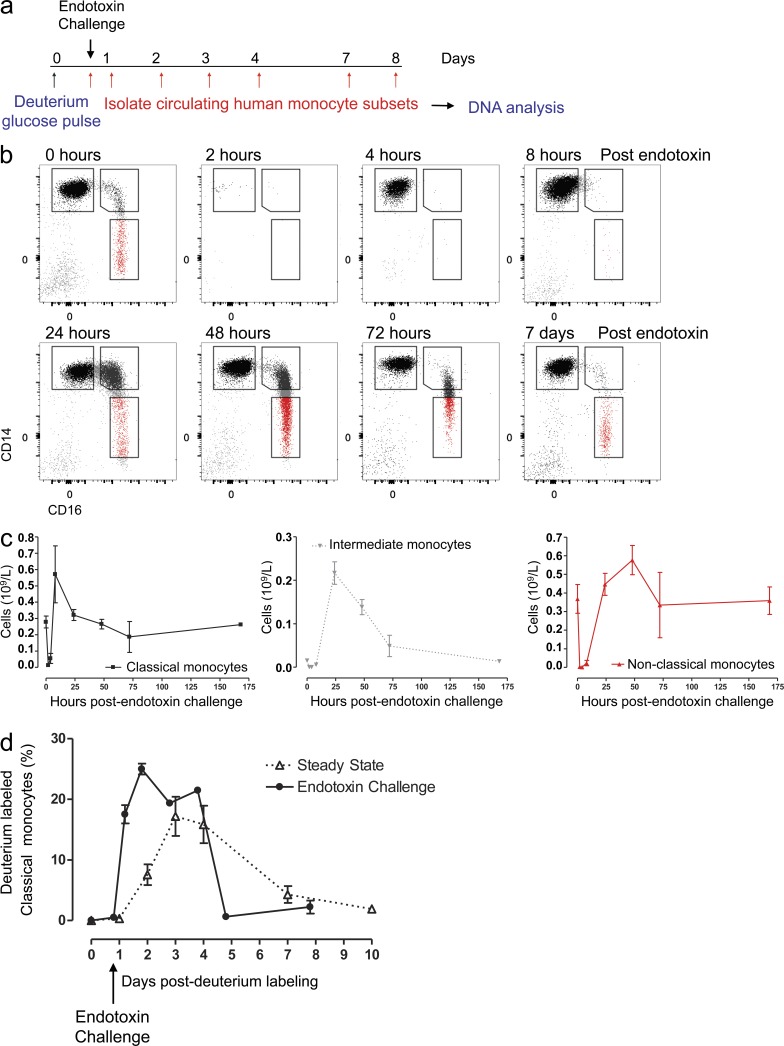
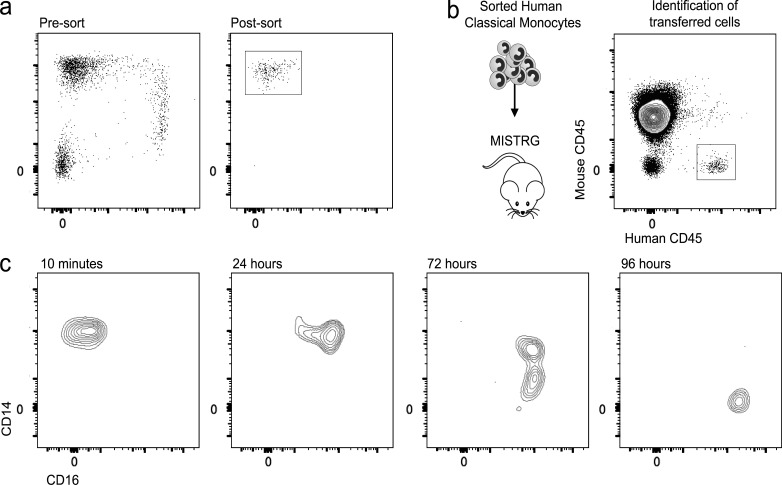
In humans, the monocyte pool comprises three subsets (classical, intermediate, and nonclassical) that circulate in dynamic equilibrium. The kinetics underlying their generation, differentiation, and disappearance are critical to understanding both steady-state homeostasis and inflammatory responses. Here, using human in vivo deuterium labeling, we demonstrate that classical monocytes emerge first from marrow, after a postmitotic interval of 1.6 d, and circulate for a day. Subsequent labeling of intermediate and nonclassical monocytes is consistent with a model of sequential transition. Intermediate and nonclassical monocytes have longer circulating lifespans (∼4 and ∼7 d, respectively). In a human experimental endotoxemia model, a transient but profound monocytopenia was observed; restoration of circulating monocytes was achieved by the early release of classical monocytes from bone marrow. The sequence of repopulation recapitulated the order of maturation in healthy homeostasis. This developmental relationship between monocyte subsets was verified by fate mapping grafted human classical monocytes into humanized mice, which were able to differentiate sequentially into intermediate and nonclassical cells.
© 2017 Patel et al.
Figures
References
-
- Arnold L., Henry A., Poron F., Baba-Amer Y., van Rooijen N., Plonquet A., Gherardi R.K., and Chazaud B.. 2007. Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J. Exp. Med. 204:1057–1069. 10.1084/jem.20070075 - DOI - PMC - PubMed
-
- Avraham-Davidi I., Yona S., Grunewald M., Landsman L., Cochain C., Silvestre J.S., Mizrahi H., Faroja M., Strauss-Ayali D., Mack M., et al. 2013. On-site education of VEGF-recruited monocytes improves their performance as angiogenic and arteriogenic accessory cells. J. Exp. Med. 210:2611–2625. 10.1084/jem.20120690 - DOI - PMC - PubMed
-
- Bain C.C., Bravo-Blas A., Scott C.L., Gomez Perdiguero E., Geissmann F., Henri S., Malissen B., Osborne L.C., Artis D., and Mowat A.M.. 2014. Constant replenishment from circulating monocytes maintains the macrophage pool in the intestine of adult mice. Nat. Immunol. 15:929–937. 10.1038/ni.2967 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
