

Ca2+-induced movement of tropomyosin on native cardiac thin filaments revealed by cryoelectron microscopy
- PMID: 28607071
- PMCID: PMC5495243
- DOI: 10.1073/pnas.1700868114
Ca2+-induced movement of tropomyosin on native cardiac thin filaments revealed by cryoelectron microscopy
Abstract
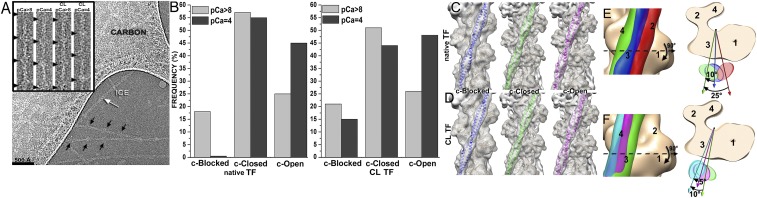
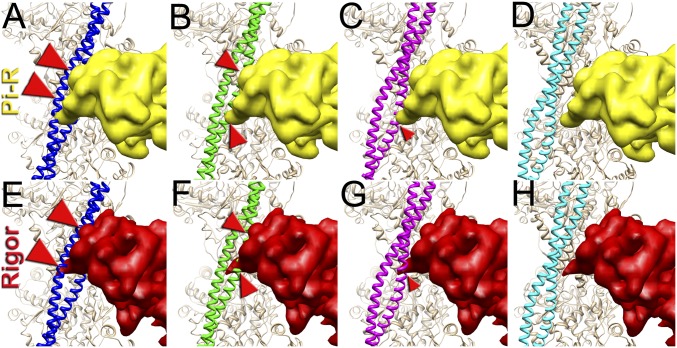
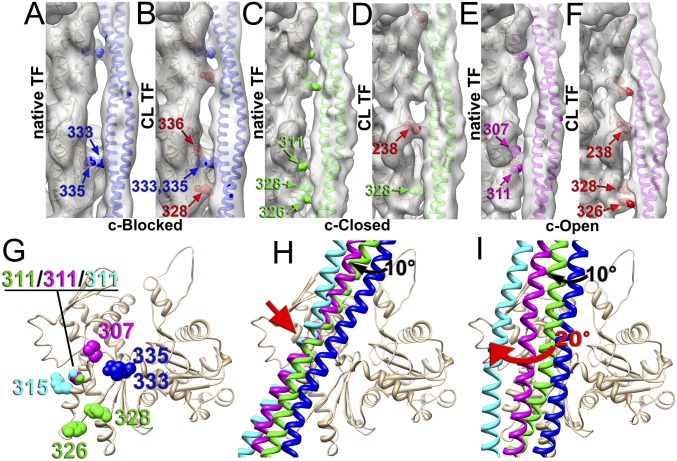
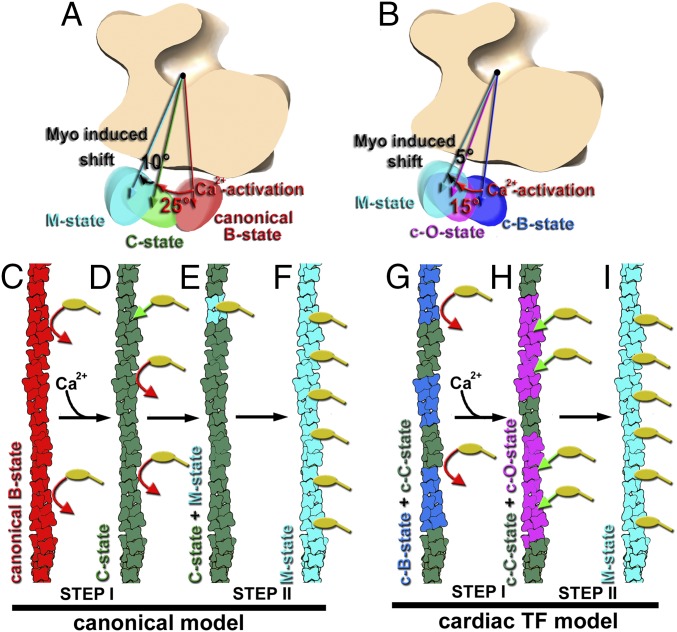
Muscle contraction relies on the interaction of myosin motors with F-actin, which is regulated through a translocation of tropomyosin by the troponin complex in response to Ca2+ The current model of muscle regulation holds that at relaxing (low-Ca2+) conditions tropomyosin blocks myosin binding sites on F-actin, whereas at activating (high-Ca2+) conditions tropomyosin translocation only partially exposes myosin binding sites on F-actin so that binding of rigor myosin is required to fully activate the thin filament (TF). Here we used a single-particle approach to helical reconstruction of frozen hydrated native cardiac TFs under relaxing and activating conditions to reveal the azimuthal movement of the tropomyosin on the surface of the native cardiac TF upon Ca2+ activation. We demonstrate that at either relaxing or activating conditions tropomyosin is not constrained in one structural state, but rather is distributed between three structural positions on the surface of the TF. We show that two of these tropomyosin positions restrain actomyosin interactions, whereas in the third position, which is significantly enhanced at high Ca2+, tropomyosin does not block myosin binding sites on F-actin. Our data provide a structural framework for the enhanced activation of the cardiac TF over the skeletal TF by Ca2+ and lead to a mechanistic model for the regulation of the cardiac TF.
Keywords: cardiac muscle regulation; cryoelectron microscopy; thin filament.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Brown JH, Cohen C. Regulation of muscle contraction by tropomyosin and troponin: How structure illuminates function. Adv Protein Chem. 2005;71:121–159. - PubMed
-
- Spudich JA, Huxley HE, Finch JT. Regulation of skeletal muscle contraction. II. Structural studies of the interaction of the tropomyosin-troponin complex with actin. J Mol Biol. 1972;72:619–632. - PubMed
-
- Vibert P, Craig R, Lehman W. Steric-model for activation of muscle thin filaments. J Mol Biol. 1997;266:8–14. - PubMed
-
- Pirani A, et al. Single particle analysis of relaxed and activated muscle thin filaments. J Mol Biol. 2005;346:761–772. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
