

Weak protein-protein interactions in live cells are quantified by cell-volume modulation
- PMID: 28607089
- PMCID: PMC5495242
- DOI: 10.1073/pnas.1700818114
Weak protein-protein interactions in live cells are quantified by cell-volume modulation
Abstract
Weakly bound protein complexes play a crucial role in metabolic, regulatory, and signaling pathways, due in part to the high tunability of their bound and unbound populations. This tunability makes weak binding (micromolar to millimolar dissociation constants) difficult to quantify under biologically relevant conditions. Here, we use rapid perturbation of cell volume to modulate the concentration of weakly bound protein complexes, allowing us to detect their dissociation constant and stoichiometry directly inside the cell. We control cell volume by modulating media osmotic pressure and observe the resulting complex association and dissociation by FRET microscopy. We quantitatively examine the interaction between GAPDH and PGK, two sequential enzymes in the glycolysis catalytic cycle. GAPDH and PGK have been shown to interact weakly, but the interaction has not been quantified in vivo. A quantitative model fits our experimental results with log Kd = -9.7 ± 0.3 and a 2:1 prevalent stoichiometry of the GAPDH:PGK complex. Cellular volume perturbation is a widely applicable tool to detect transient protein interactions and other biomolecular interactions in situ. Our results also suggest that cells could use volume change (e.g., as occurs upon entry to mitosis) to regulate function by altering biomolecular complex concentrations.
Keywords: FRET; cell volume; live-cell microscopy; protein–protein interactions; quinary interactions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
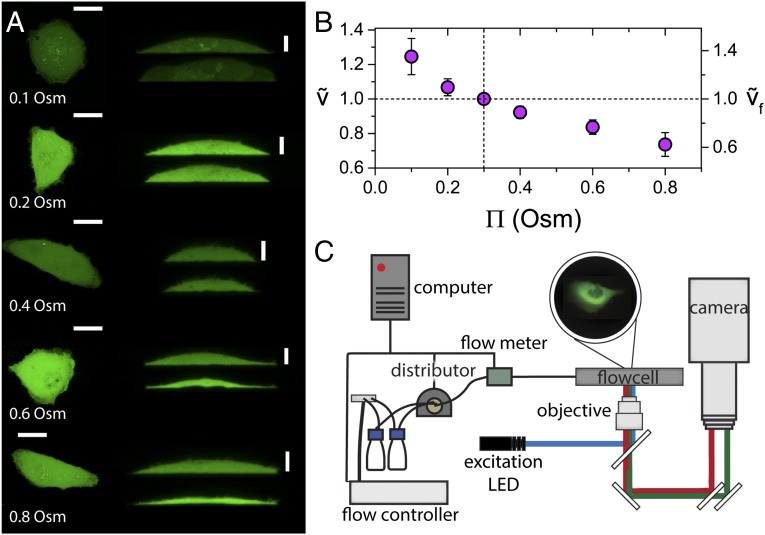
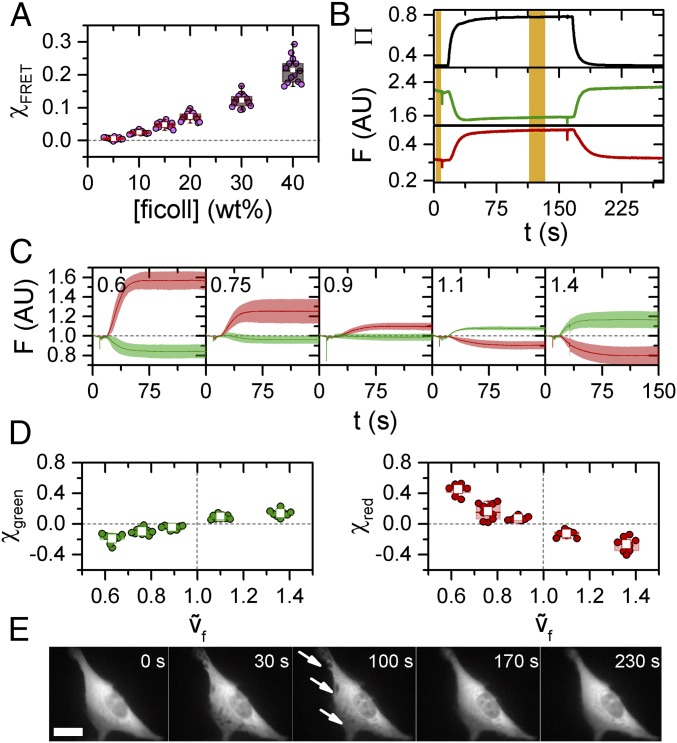
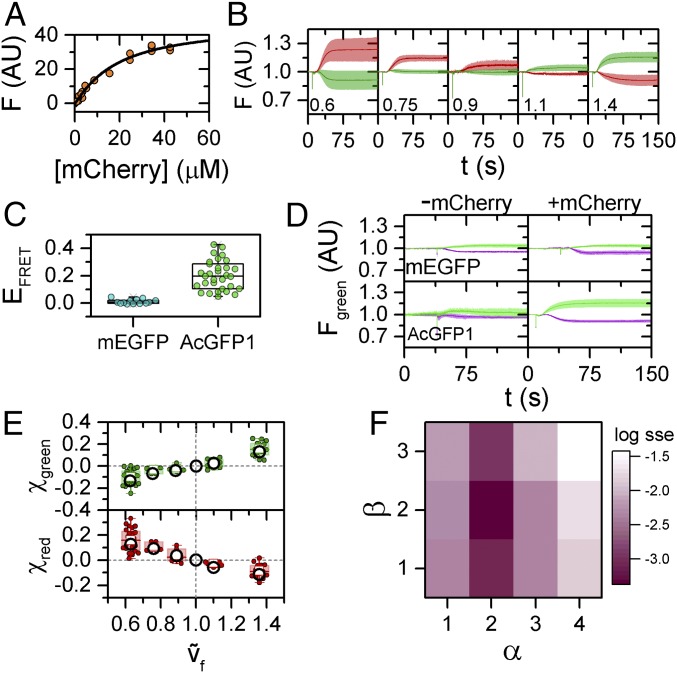
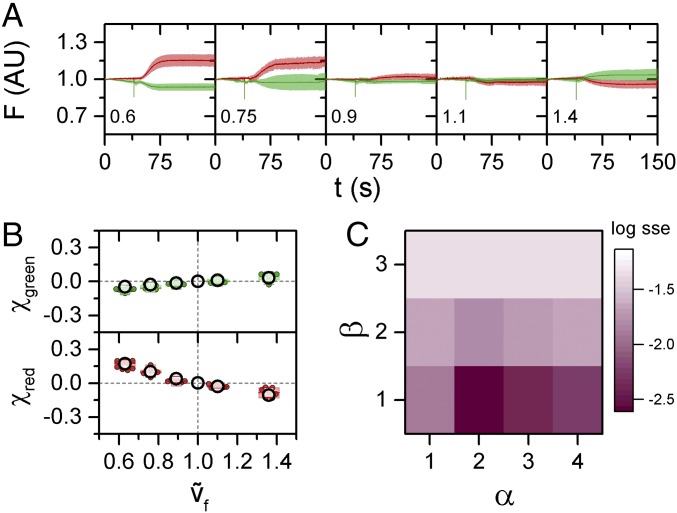
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
