

Epigenetic Transcriptional Memory of GAL Genes Depends on Growth in Glucose and the Tup1 Transcription Factor in Saccharomyces cerevisiae
- PMID: 28607146
- PMCID: PMC5560796
- DOI: 10.1534/genetics.117.201632
Epigenetic Transcriptional Memory of GAL Genes Depends on Growth in Glucose and the Tup1 Transcription Factor in Saccharomyces cerevisiae
Abstract
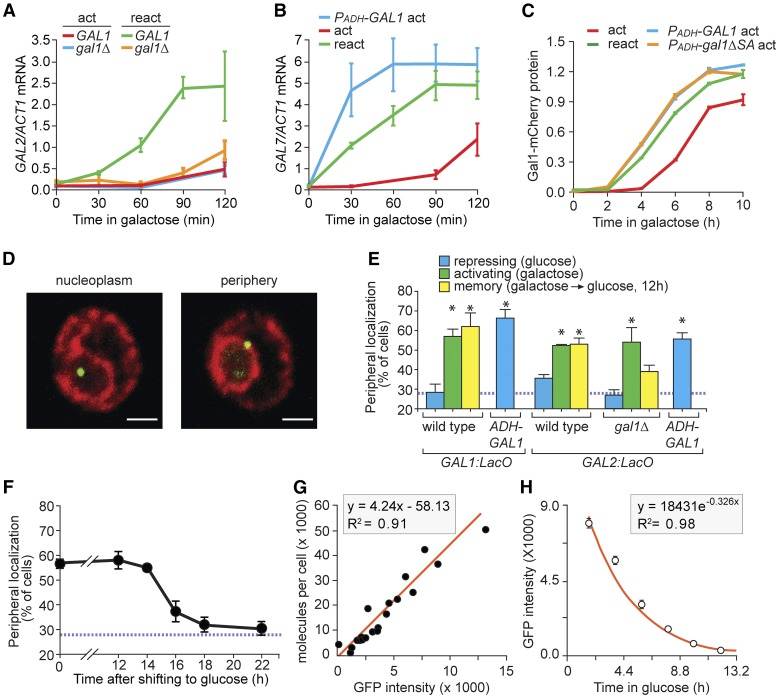
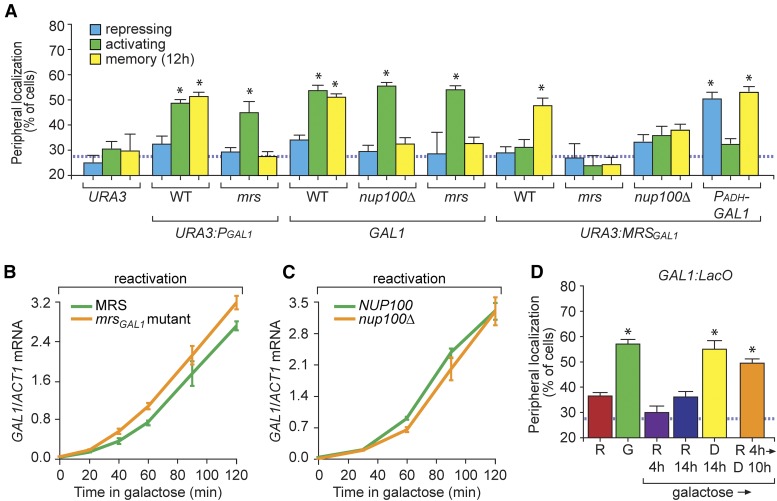
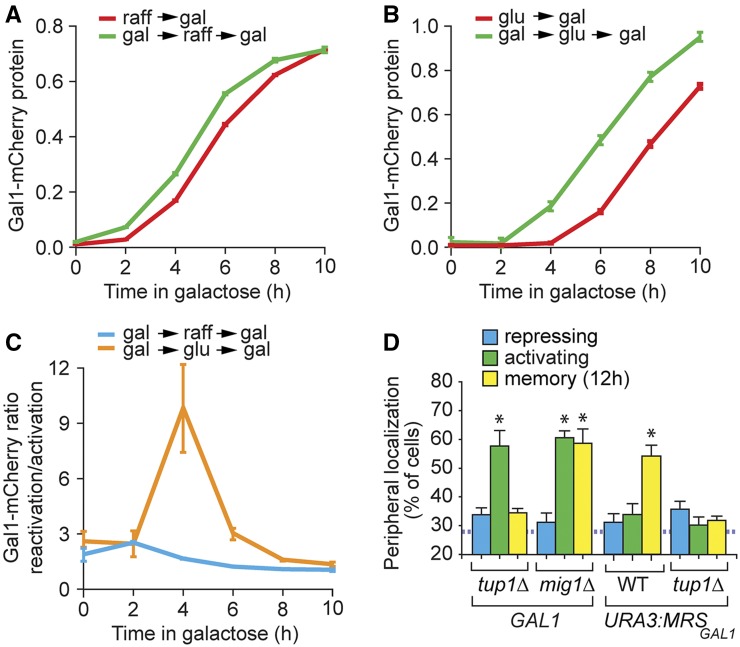
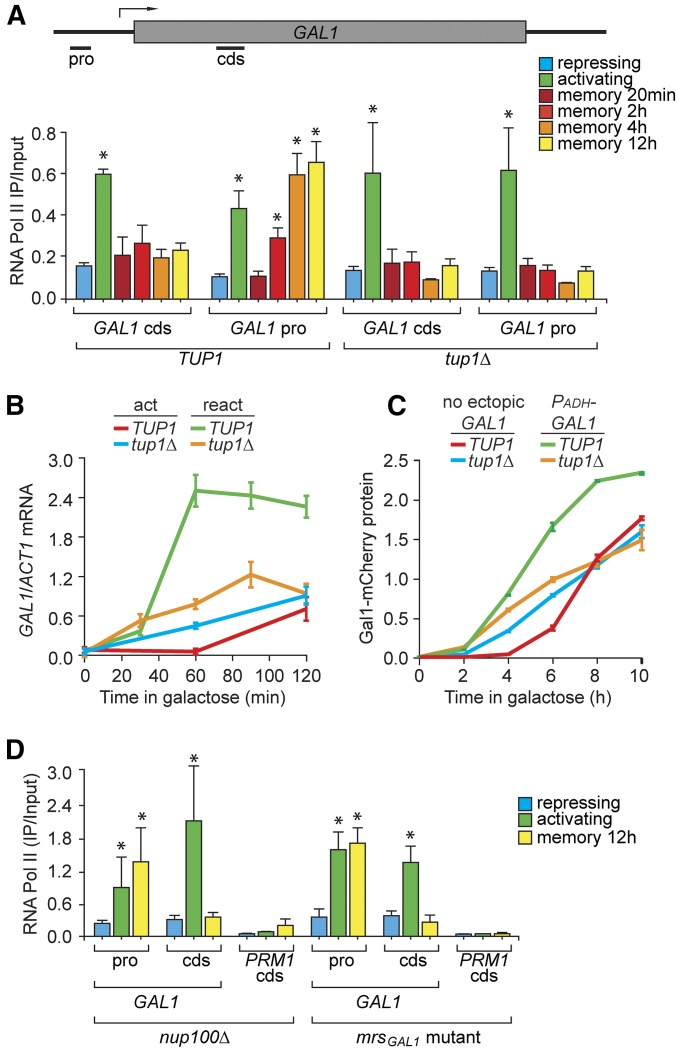
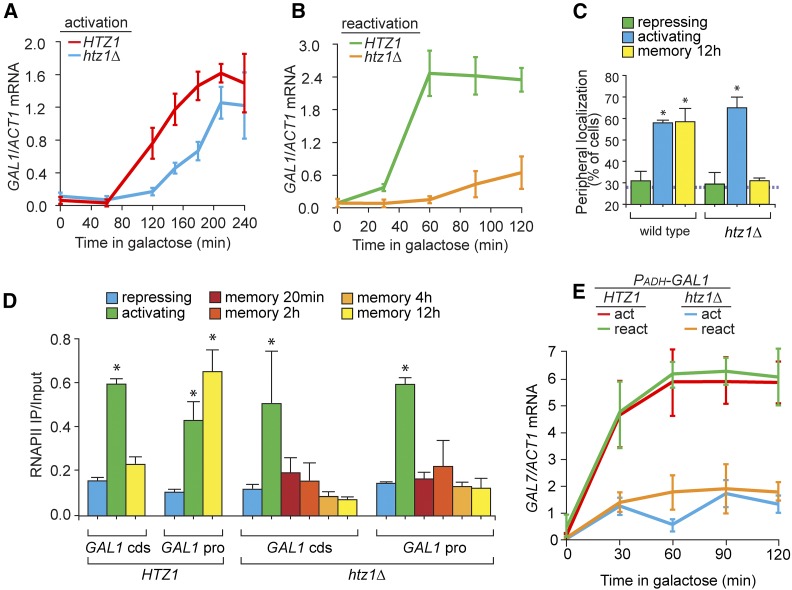
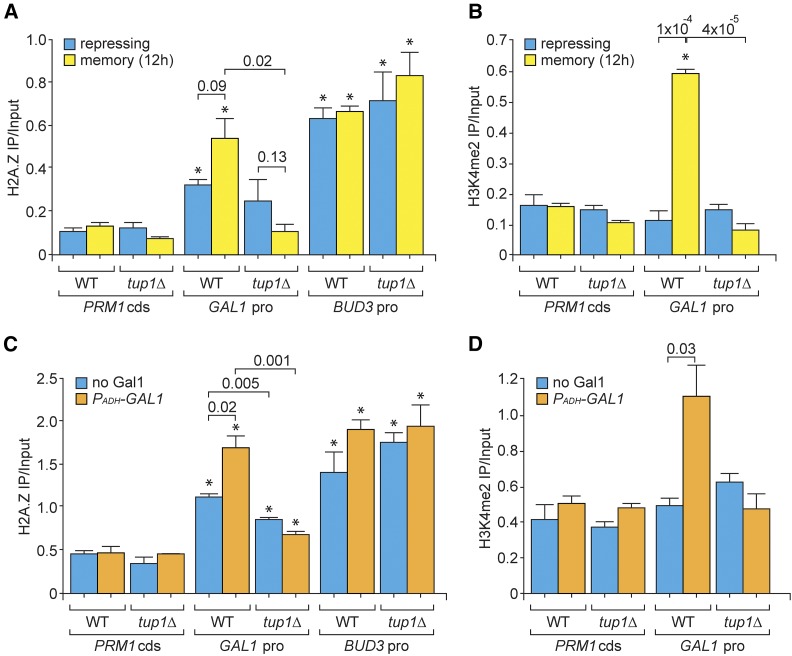
Previously expressed inducible genes can remain poised for faster reactivation for multiple cell divisions, a conserved phenomenon called epigenetic transcriptional memory. The GAL genes in Saccharomyces cerevisiae show faster reactivation for up to seven generations after being repressed. During memory, previously produced Gal1 protein enhances the rate of reactivation of GAL1, GAL10, GAL2, and GAL7 These genes also interact with the nuclear pore complex (NPC) and localize to the nuclear periphery both when active and during memory. Peripheral localization of GAL1 during memory requires the Gal1 protein, a memory-specific cis-acting element in the promoter, and the NPC protein Nup100 However, unlike other examples of transcriptional memory, the interaction with NPC is not required for faster GAL gene reactivation. Rather, downstream of Gal1, the Tup1 transcription factor and growth in glucose promote GAL transcriptional memory. Cells only show signs of memory and only benefit from memory when growing in glucose. Tup1 promotes memory-specific chromatin changes at the GAL1 promoter: incorporation of histone variant H2A.Z and dimethylation of histone H3, lysine 4. Tup1 and H2A.Z function downstream of Gal1 to promote binding of a preinitiation form of RNA Polymerase II at the GAL1 promoter, poising the gene for faster reactivation. This mechanism allows cells to integrate a previous experience (growth in galactose, reflected by Gal1 levels) with current conditions (growth in glucose, potentially through Tup1 function) to overcome repression and to poise critical GAL genes for future reactivation.
Keywords: GAL genes; RNA polymerase II; chromatin; epigenetic; gene positioning; nuclear pore complex; transcriptional memory.
Copyright © 2017 by the Genetics Society of America.
Figures
Similar articles
-
The Tup1 corepressor directs Htz1 deposition at a specific promoter nucleosome marking the GAL1 gene for rapid activation.Mol Cell Biol. 2007 Jun;27(11):4198-205. doi: 10.1128/MCB.00238-07. Epub 2007 Mar 26. Mol Cell Biol. 2007. PMID: 17387147 Free PMC article.
-
The nuclear pore regulates GAL1 gene transcription by controlling the localization of the SUMO protease Ulp1.Mol Cell. 2013 Sep 26;51(6):807-18. doi: 10.1016/j.molcel.2013.08.047. Mol Cell. 2013. PMID: 24074957
-
H2A.Z-mediated localization of genes at the nuclear periphery confers epigenetic memory of previous transcriptional state.PLoS Biol. 2007 Apr;5(4):e81. doi: 10.1371/journal.pbio.0050081. PLoS Biol. 2007. PMID: 17373856 Free PMC article.
-
Transcriptional memory at the nuclear periphery.Curr Opin Cell Biol. 2009 Feb;21(1):127-33. doi: 10.1016/j.ceb.2009.01.007. Epub 2009 Jan 30. Curr Opin Cell Biol. 2009. PMID: 19181512 Free PMC article. Review.
-
Epigenetics of the yeast galactose genetic switch.J Biosci. 2009 Oct;34(4):513-22. doi: 10.1007/s12038-009-0070-y. J Biosci. 2009. PMID: 19920337 Review.
Cited by
-
Histone Methylation and Memory of Environmental Stress.Cells. 2019 Apr 10;8(4):339. doi: 10.3390/cells8040339. Cells. 2019. PMID: 30974922 Free PMC article. Review.
-
Capturing and Understanding the Dynamics and Heterogeneity of Gene Expression in the Living Cell.Int J Mol Sci. 2020 Nov 5;21(21):8278. doi: 10.3390/ijms21218278. Int J Mol Sci. 2020. PMID: 33167354 Free PMC article. Review.
-
HapX-mediated H2B deub1 and SreA-mediated H2A.Z deposition coordinate in fungal iron resistance.Nucleic Acids Res. 2023 Oct 27;51(19):10238-10260. doi: 10.1093/nar/gkad708. Nucleic Acids Res. 2023. PMID: 37650633 Free PMC article.
-
Plant defense compound triggers mycotoxin synthesis by regulating H2B ub1 and H3K4 me2/3 deposition.New Phytol. 2021 Dec;232(5):2106-2123. doi: 10.1111/nph.17718. Epub 2021 Sep 30. New Phytol. 2021. PMID: 34480757 Free PMC article.
-
AP-1 Mediates Cellular Adaptation and Memory Formation During Therapy Resistance.bioRxiv [Preprint]. 2024 Jul 25:2024.07.25.604999. doi: 10.1101/2024.07.25.604999. bioRxiv. 2024. PMID: 39091739 Free PMC article. Preprint.
References
-
- Brickner D. G., Light W., Brickner J. H., 2010. Quantitative localization of chromosomal loci by immunofluorescence. Methods Enzymol. 470: 569–580. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
