

A retinal code for motion along the gravitational and body axes
- PMID: 28607486
- PMCID: PMC5729591
- DOI: 10.1038/nature22818
A retinal code for motion along the gravitational and body axes
Abstract
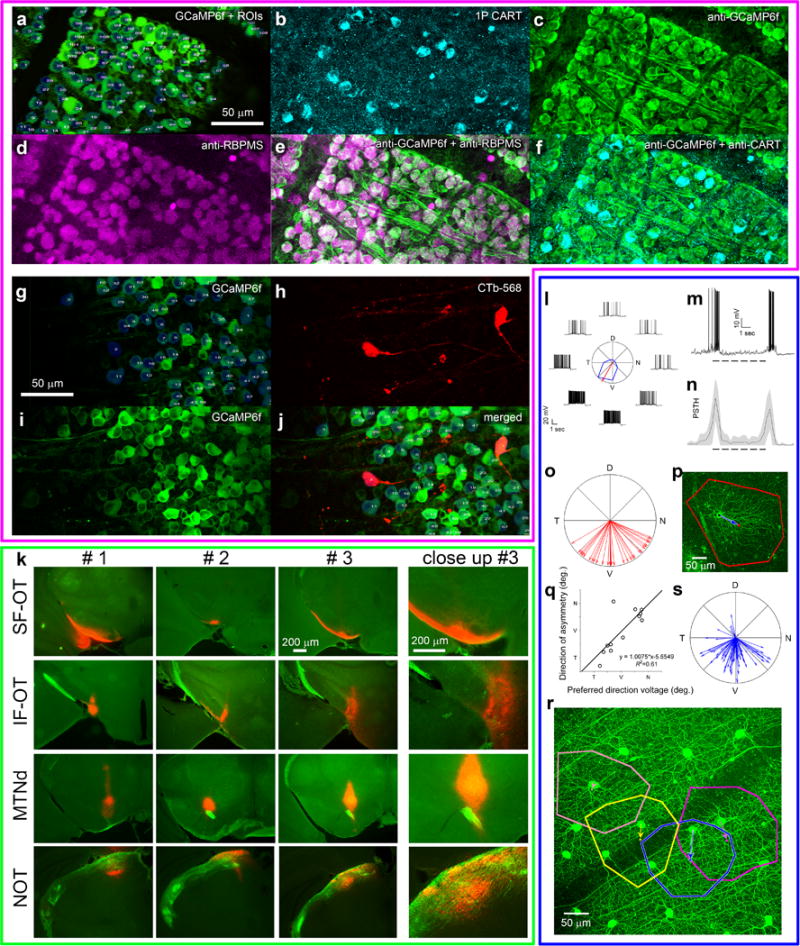
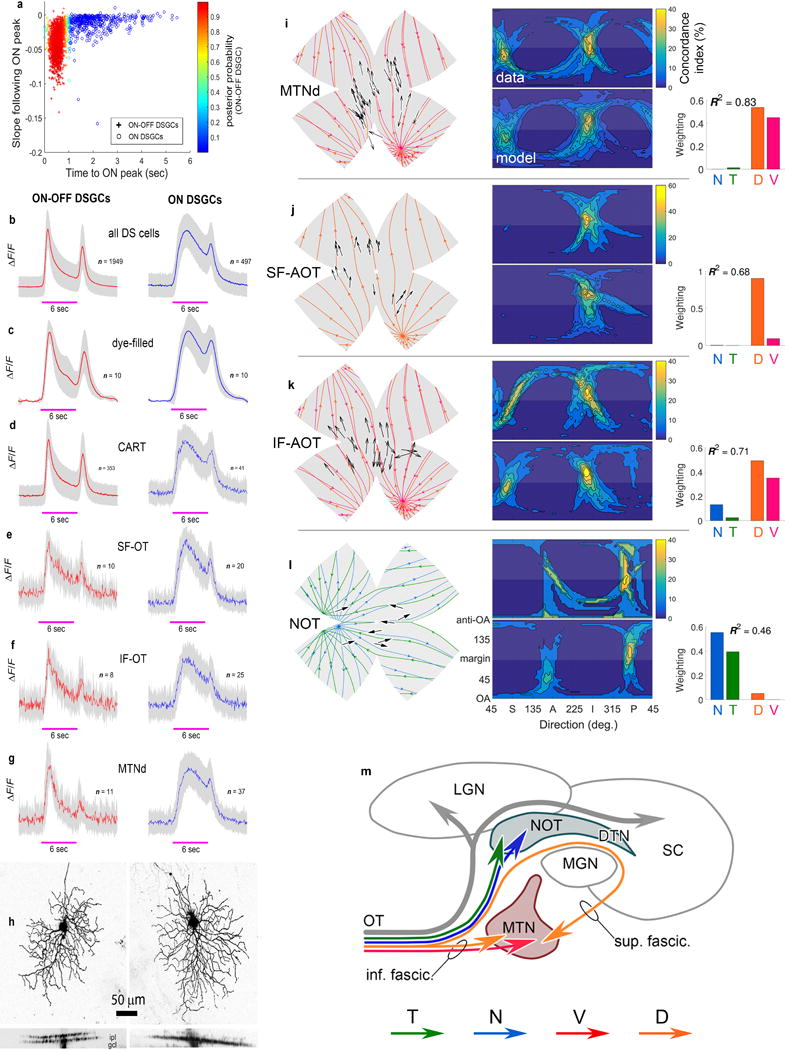
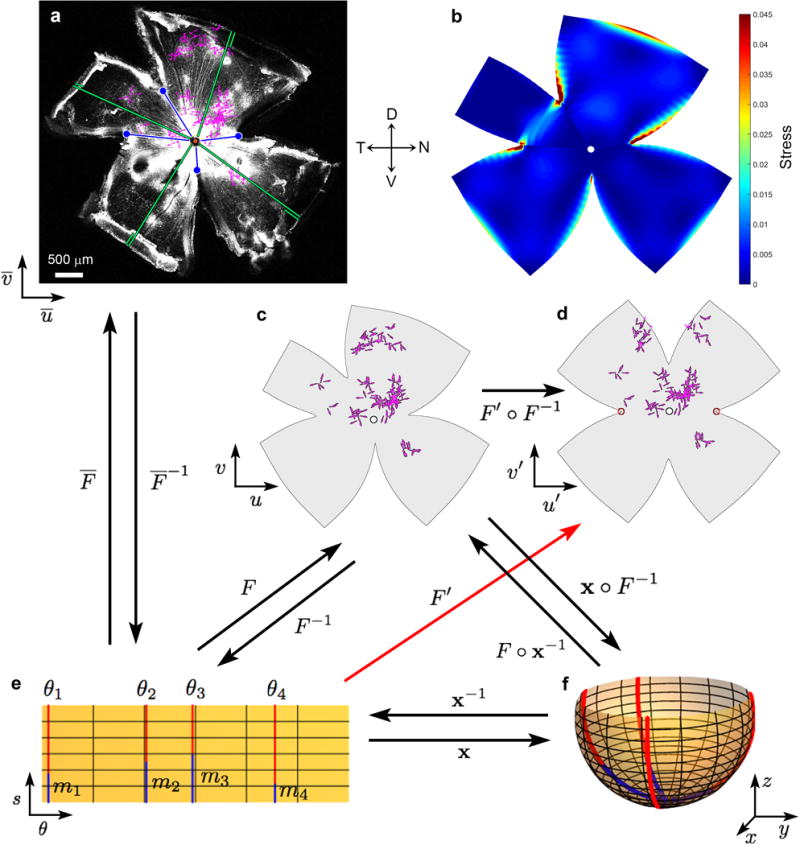
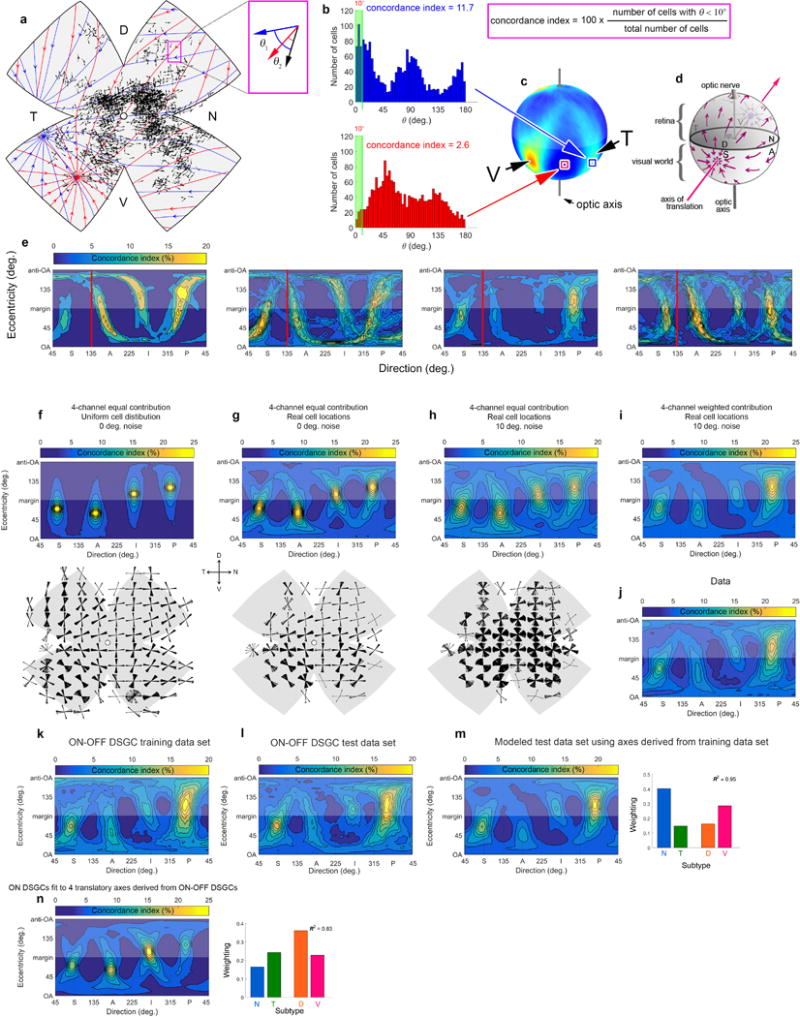
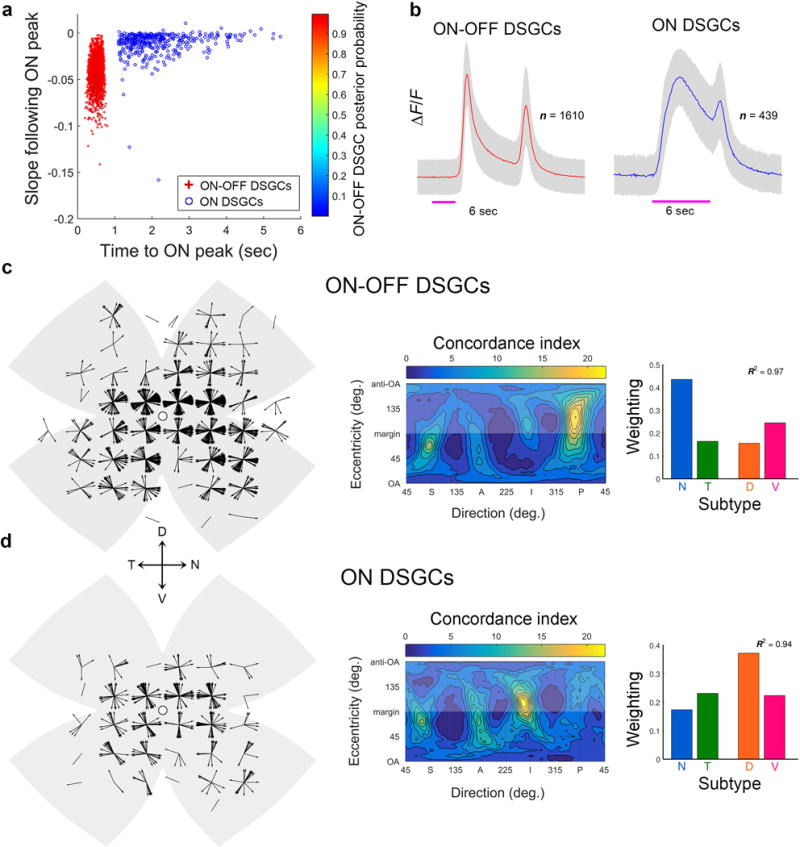
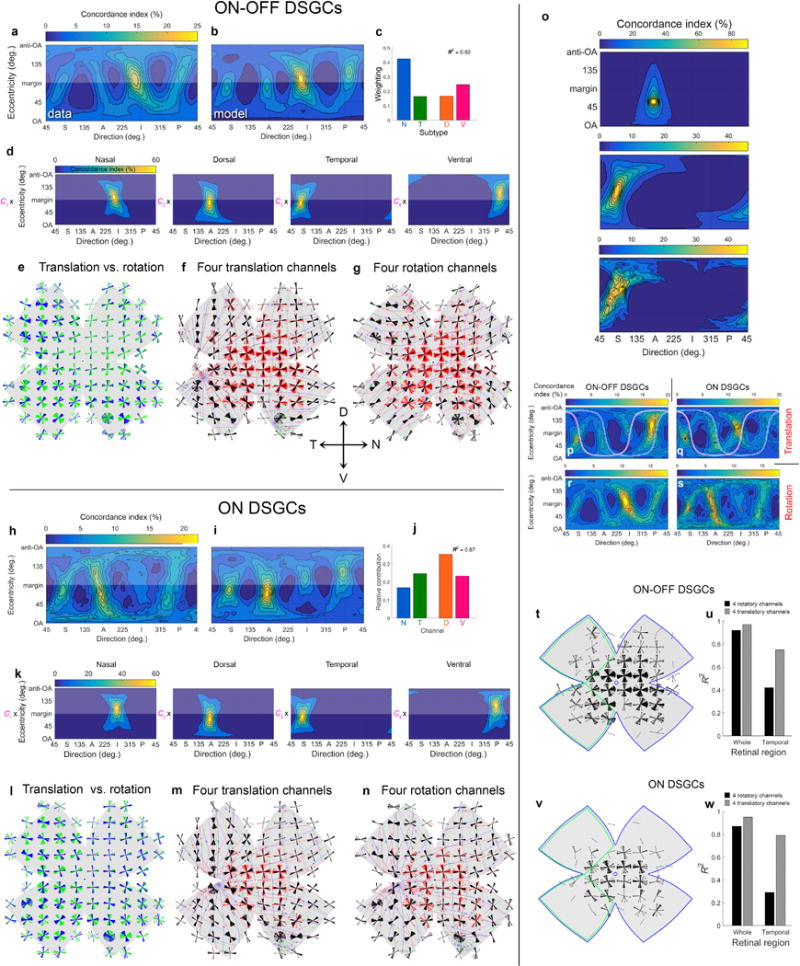
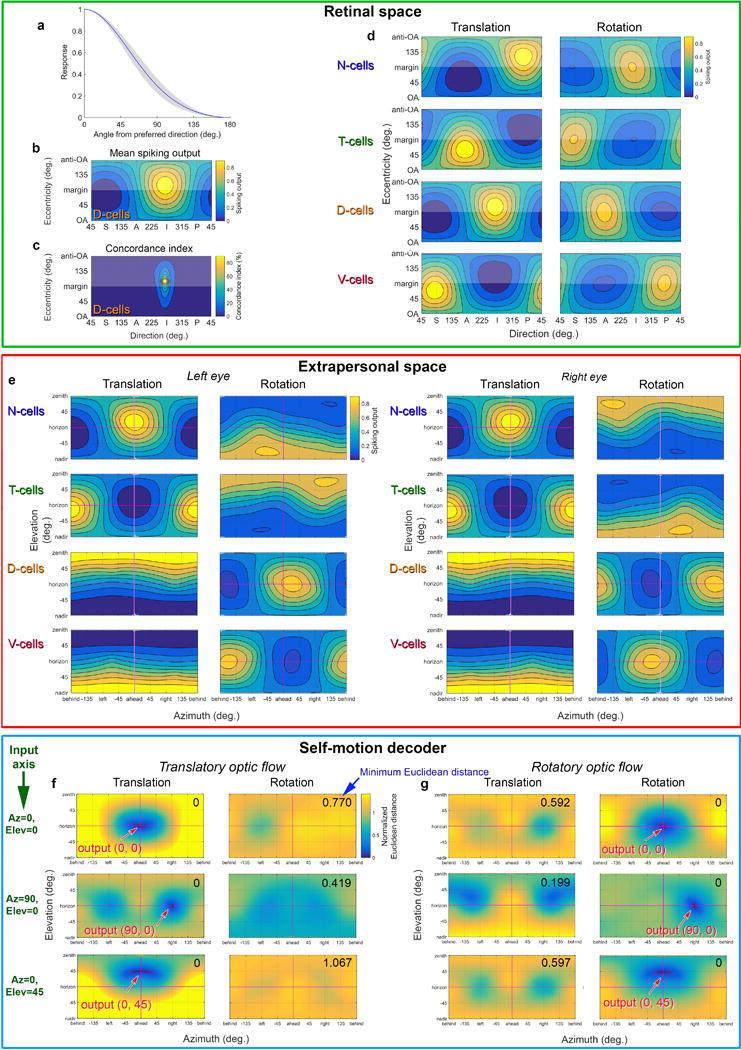
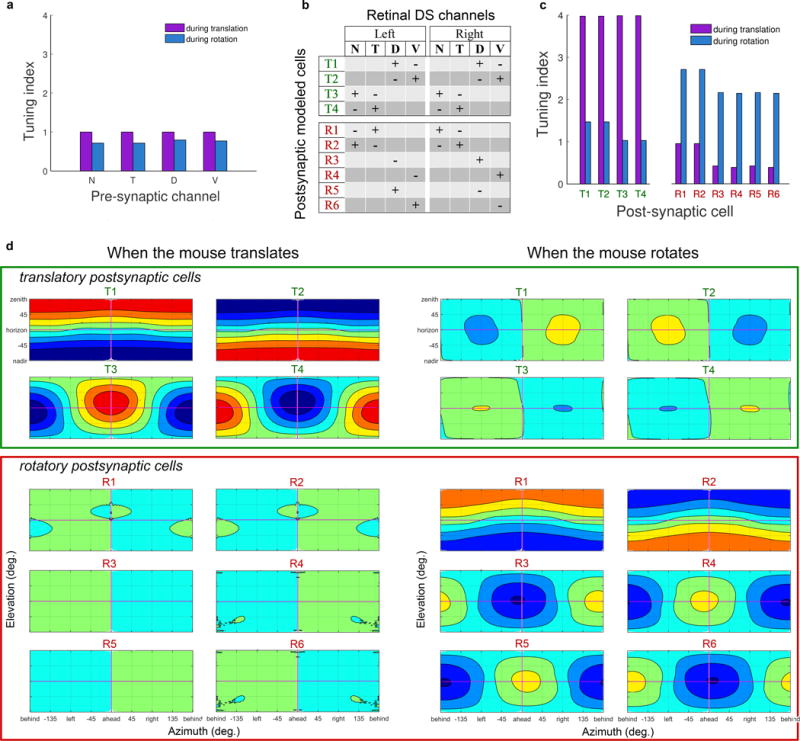
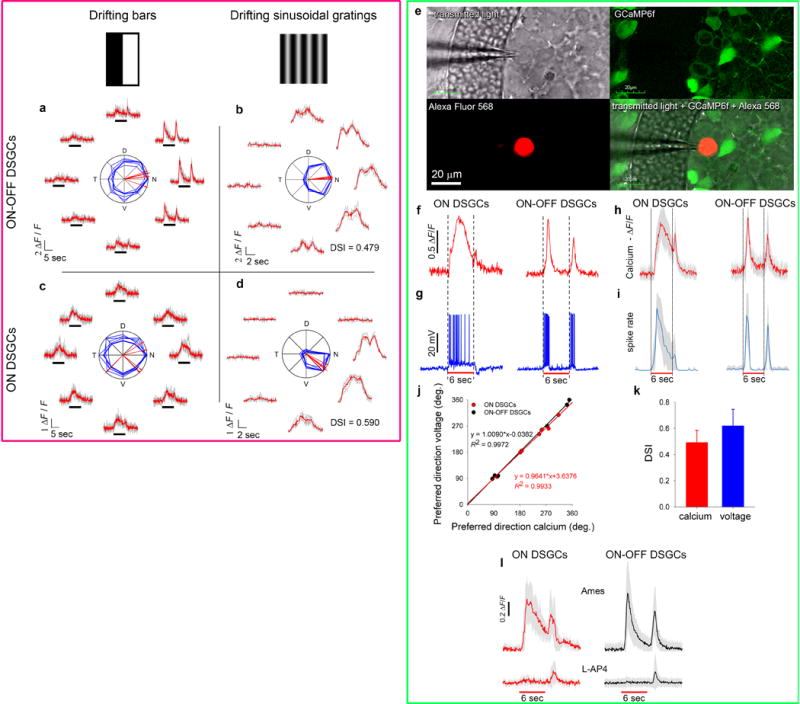
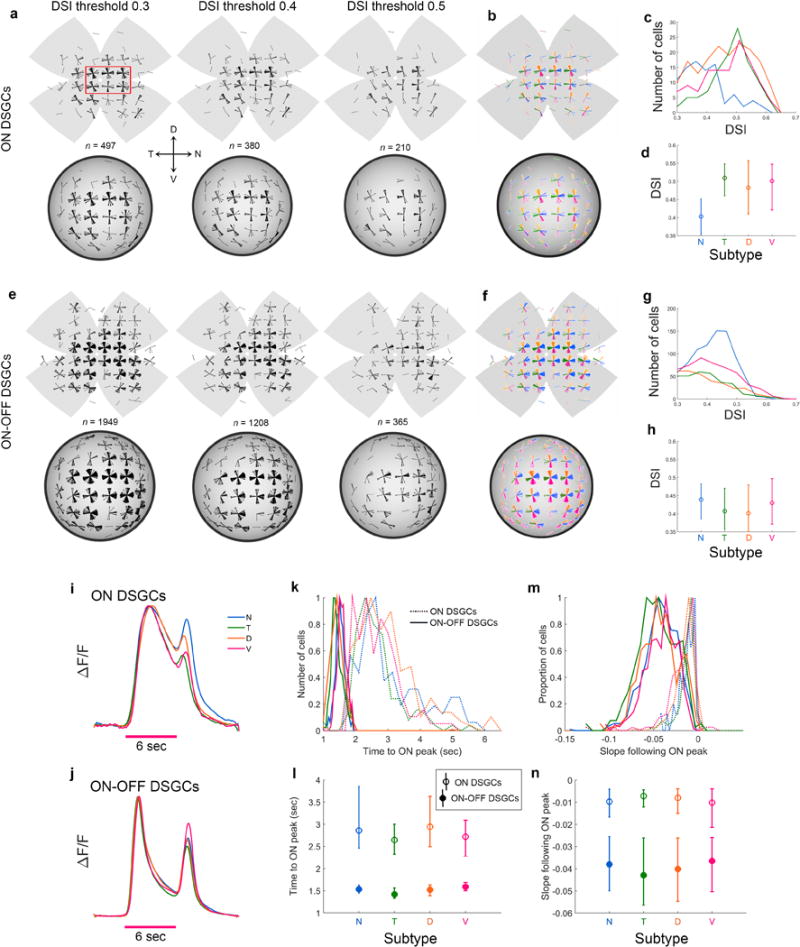
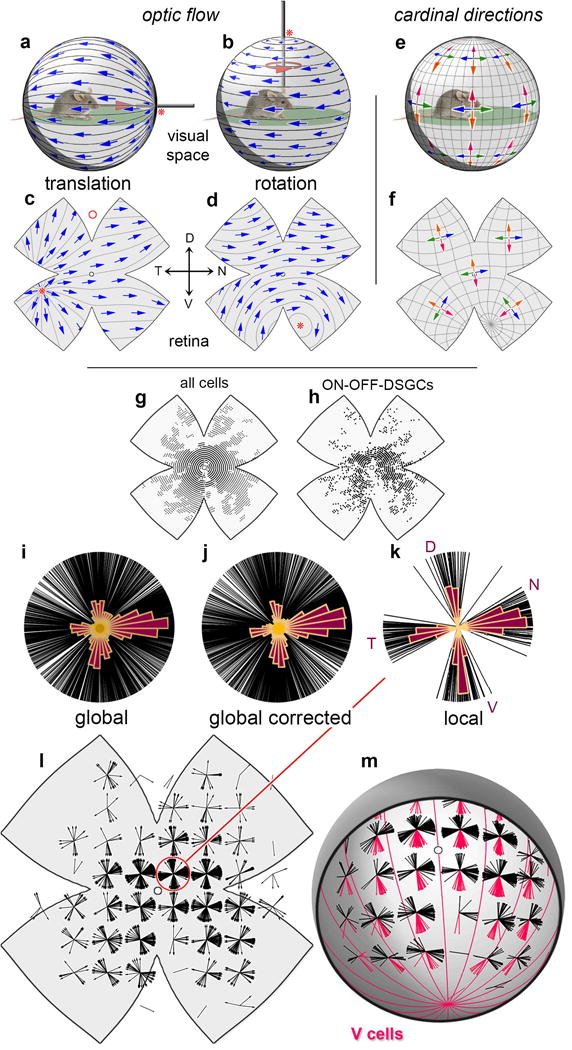
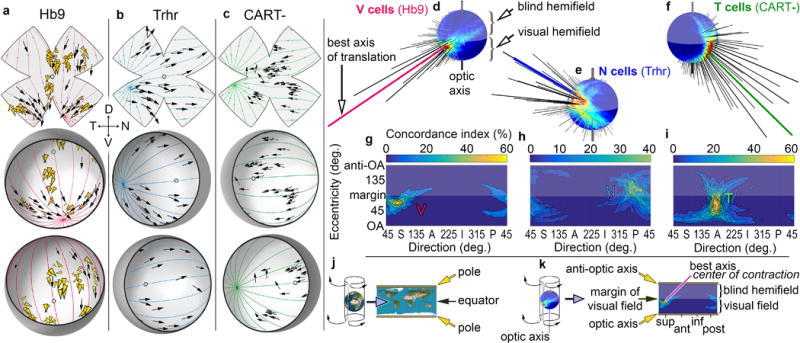
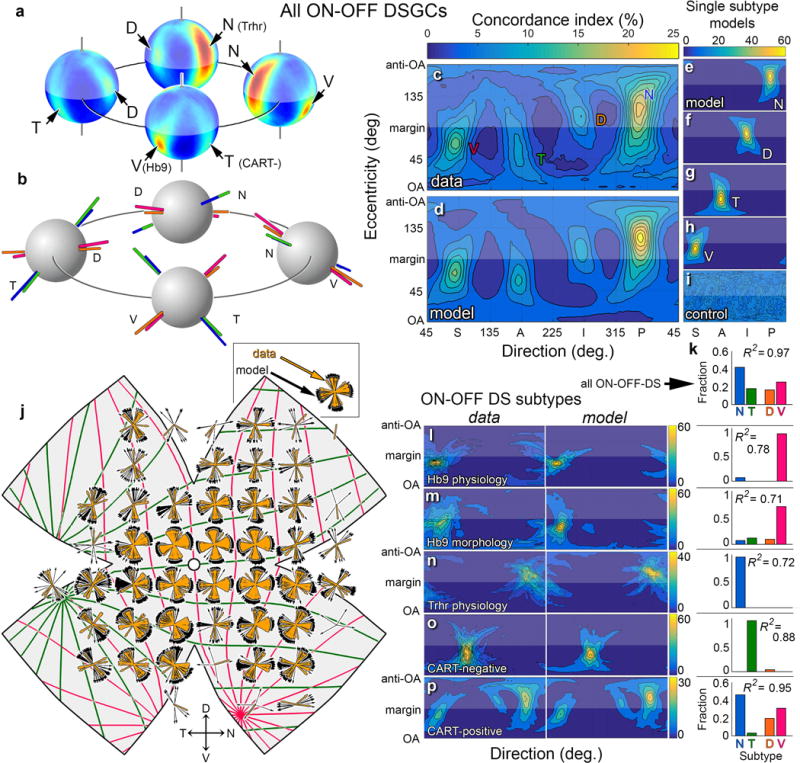
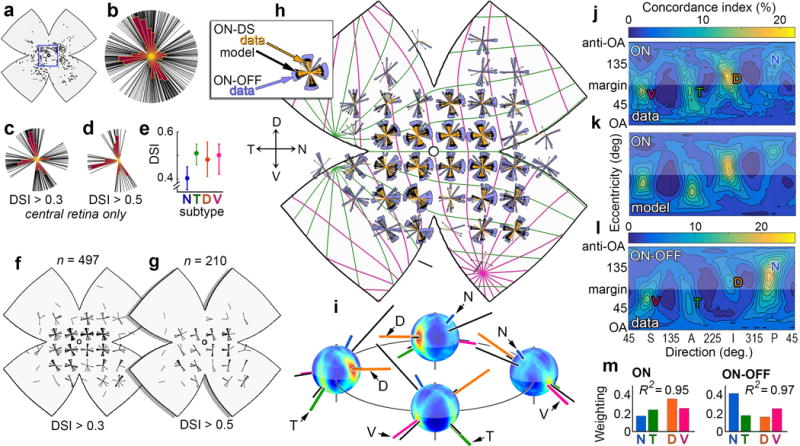
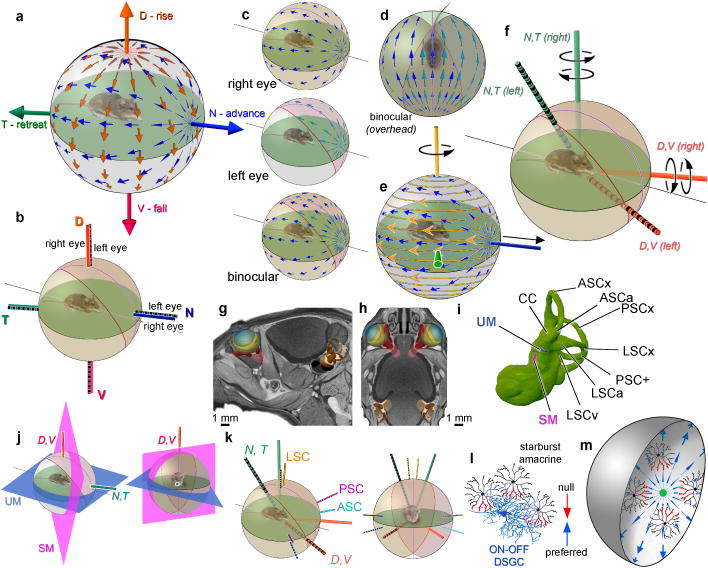
Self-motion triggers complementary visual and vestibular reflexes supporting image-stabilization and balance. Translation through space produces one global pattern of retinal image motion (optic flow), rotation another. We examined the direction preferences of direction-sensitive ganglion cells (DSGCs) in flattened mouse retinas in vitro. Here we show that for each subtype of DSGC, direction preference varies topographically so as to align with specific translatory optic flow fields, creating a neural ensemble tuned for a specific direction of motion through space. Four cardinal translatory directions are represented, aligned with two axes of high adaptive relevance: the body and gravitational axes. One subtype maximizes its output when the mouse advances, others when it retreats, rises or falls. Two classes of DSGCs, namely, ON-DSGCs and ON-OFF-DSGCs, share the same spatial geometry but weight the four channels differently. Each subtype ensemble is also tuned for rotation. The relative activation of DSGC channels uniquely encodes every translation and rotation. Although retinal and vestibular systems both encode translatory and rotatory self-motion, their coordinate systems differ.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Vision: These retinas are made for walkin'.Nature. 2017 Jun 22;546(7659):476-477. doi: 10.1038/nature22505. Epub 2017 Jun 7. Nature. 2017. PMID: 28607483 No abstract available.
References
-
- Vaney DI, He S, Taylor WR, Levick WR. In: Motion Vision - Computational, Neural, and Ecological Constraints. Zanker JM, Zeil, editors. Springer Verlag; 2001. pp. 13–56.
-
- Taylor WR, He S, Levick WR, Vaney DI. Dendritic computation of direction selectivity by retinal ganglion cells. Science. 2000;289:2347–2350. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
