

Could LC-NE-Dependent Adjustment of Neural Gain Drive Functional Brain Network Reorganization?
- PMID: 28607776
- PMCID: PMC5457760
- DOI: 10.1155/2017/4328015
Could LC-NE-Dependent Adjustment of Neural Gain Drive Functional Brain Network Reorganization?
Abstract
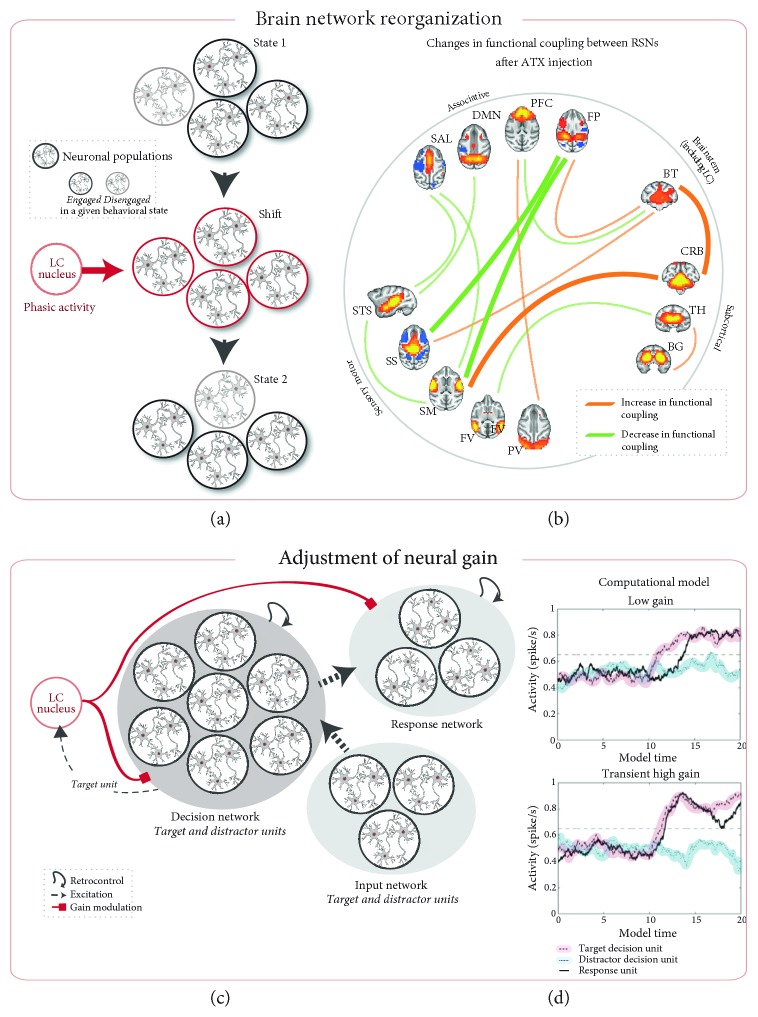
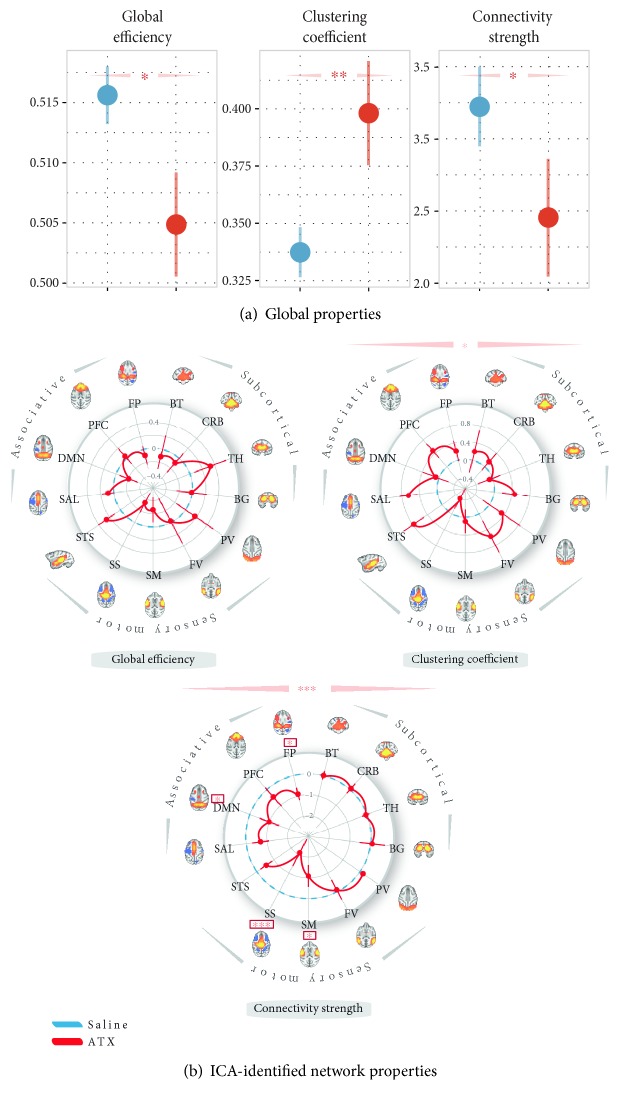
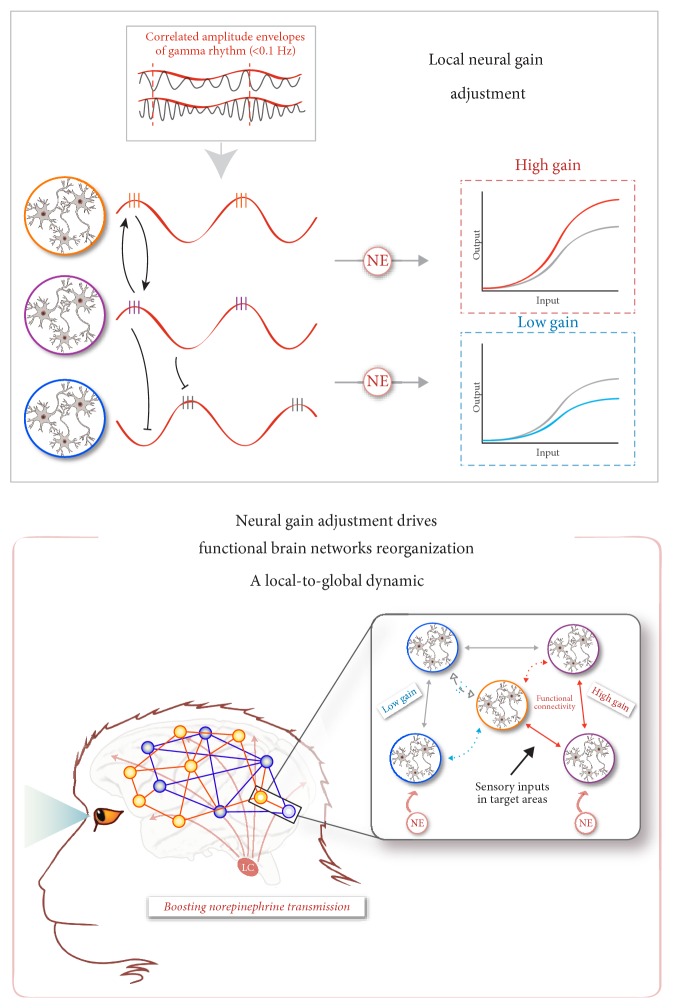
The locus coeruleus-norepinephrine (LC-NE) system is thought to act at synaptic, cellular, microcircuit, and network levels to facilitate cognitive functions through at least two different processes, not mutually exclusive. Accordingly, as a reset signal, the LC-NE system could trigger brain network reorganizations in response to salient information in the environment and/or adjust the neural gain within its target regions to optimize behavioral responses. Here, we provide evidence of the co-occurrence of these two mechanisms at the whole-brain level, in resting-state conditions following a pharmacological stimulation of the LC-NE system. We propose that these two mechanisms are interdependent such that the LC-NE-dependent adjustment of the neural gain inferred from the clustering coefficient could drive functional brain network reorganizations through coherence in the gamma rhythm. Via the temporal dynamic of gamma-range band-limited power, the release of NE could adjust the neural gain, promoting interactions only within the neuronal populations whose amplitude envelopes are correlated, thus making it possible to reorganize neuronal ensembles, functional networks, and ultimately, behavioral responses. Thus, our proposal offers a unified framework integrating the putative influence of the LC-NE system on both local- and long-range adjustments of brain dynamics underlying behavioral flexibility.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
