

Global proteomic analysis of advanced glycation end products in the Arabidopsis proteome provides evidence for age-related glycation hot spots
- PMID: 28611063
- PMCID: PMC5612108
- DOI: 10.1074/jbc.M117.794537
Global proteomic analysis of advanced glycation end products in the Arabidopsis proteome provides evidence for age-related glycation hot spots
Abstract
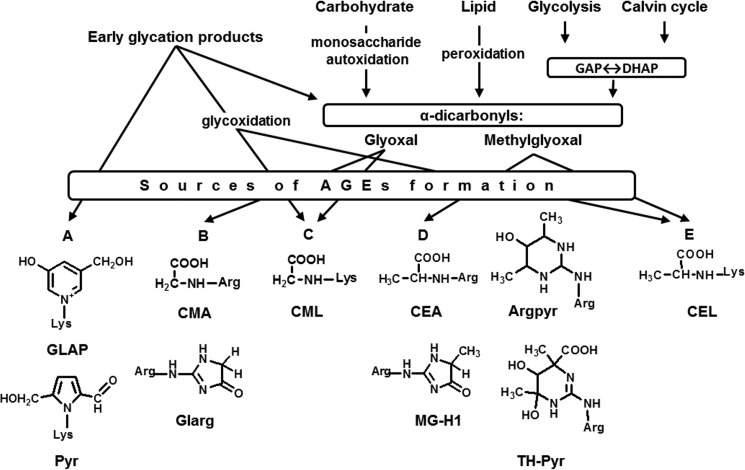
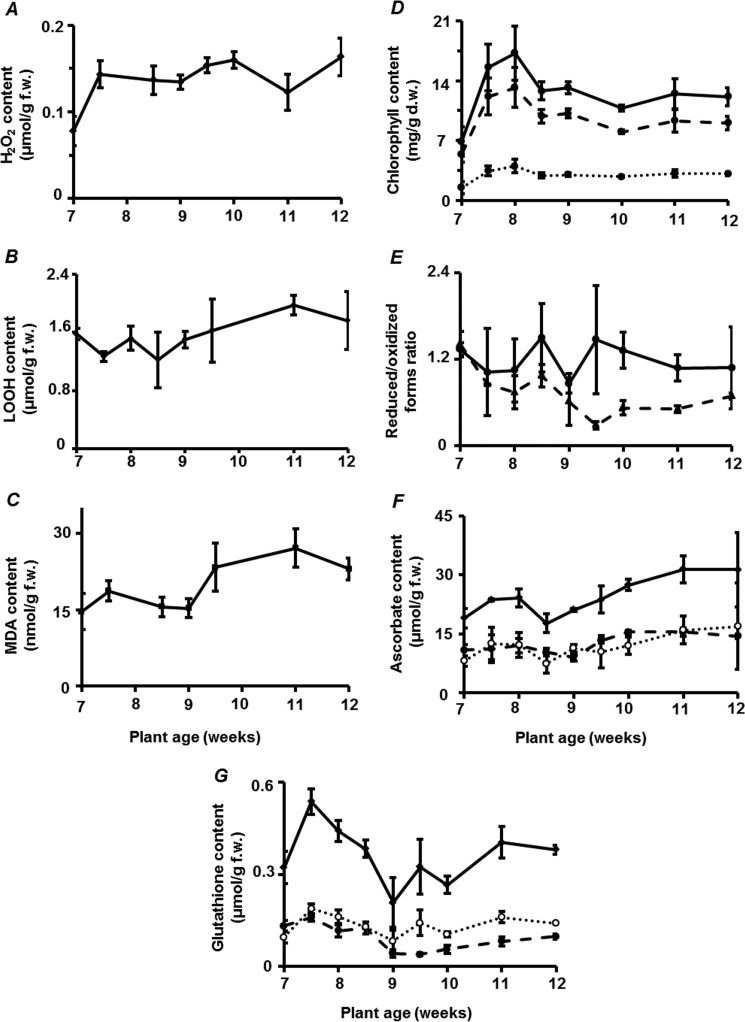
Glycation is a post-translational modification resulting from the interaction of protein amino and guanidino groups with carbonyl compounds. Initially, amino groups react with reducing carbohydrates, yielding Amadori and Heyns compounds. Their further degradation results in formation of advanced glycation end products (AGEs), also originating from α-dicarbonyl products of monosaccharide autoxidation and primary metabolism. In mammals, AGEs are continuously formed during the life of the organism, accumulate in tissues, are well-known markers of aging, and impact age-related tissue stiffening and atherosclerotic changes. However, the role of AGEs in age-related molecular alterations in plants is still unknown. To fill this gap, we present here a comprehensive study of the age-related changes in the Arabidopsis thaliana glycated proteome, including the proteins affected and specific glycation sites therein. We also consider the qualitative and quantitative changes in glycation patterns in terms of the general metabolic background, pathways of AGE formation, and the status of plant anti-oxidative/anti-glycative defense. Although the patterns of glycated proteins were only minimally influenced by plant age, the abundance of 96 AGE sites in 71 proteins was significantly affected in an age-dependent manner and clearly indicated the existence of age-related glycation hot spots in the plant proteome. Homology modeling revealed glutamyl and aspartyl residues in close proximity (less than 5 Å) to these sites in three aging-specific and eight differentially glycated proteins, four of which were modified in catalytic domains. Thus, the sites of glycation hot spots might be defined by protein structure that indicates, at least partly, site-specific character of glycation.
Keywords: Arabidopsis thaliana; advanced glycation end products (AGEs); aging; glycation; homology modeling; hot spots of glycation; metabolomics; oxidative stress; post-translational modification (PTM); proteomics.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures








References
-
- Vistoli G., De Maddis D., Cipak A., Zarkovic N., Carini M., and Aldini G. (2013) Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): an overview of their mechanisms of formation. Free Radic. Res. 47, 3–27 - PubMed
-
- Heyns K., and Beilfuss W. (1970) Ketosylamine rearrangement of d-threo-pentulose (d-xylulose) with α-amino acids. Chem. Ber. 103, 2873–2876 - PubMed
-
- Hodge J. E. (1955) The Amadori rearrangement. Adv. Carbohydr. Chem. 10, 169–205 - PubMed
-
- Greifenhagen U., Frolov A., and Hoffmann R. (2015) Oxidative degradation of N(ϵ)-fructosylamine-substituted peptides in heated aqueous systems. Amino Acids 47, 1065–1076 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
