

The Aspergillus fumigatus CrzA Transcription Factor Activates Chitin Synthase Gene Expression during the Caspofungin Paradoxical Effect
- PMID: 28611248
- PMCID: PMC5472186
- DOI: 10.1128/mBio.00705-17
The Aspergillus fumigatus CrzA Transcription Factor Activates Chitin Synthase Gene Expression during the Caspofungin Paradoxical Effect
Abstract
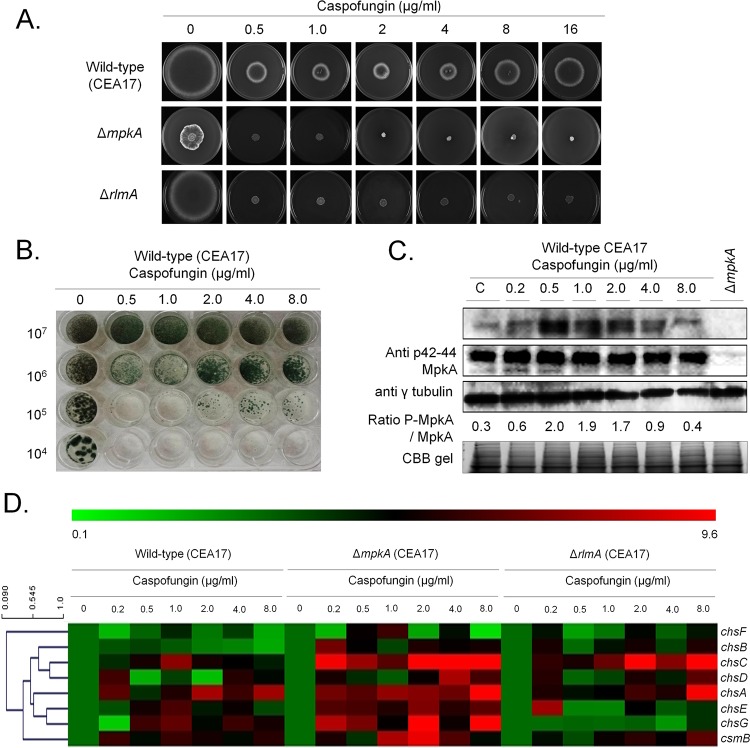
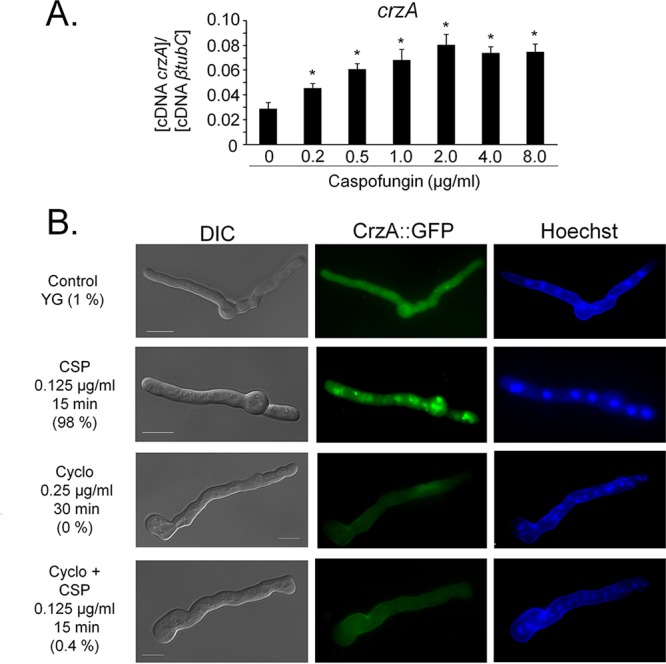
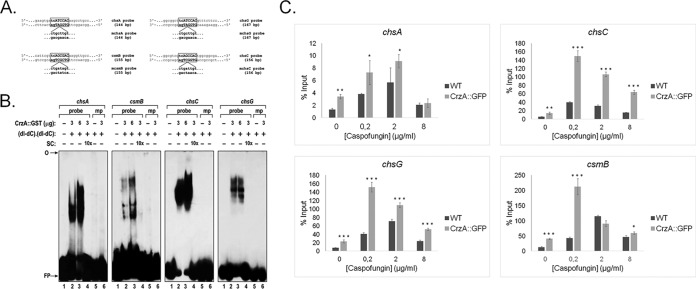
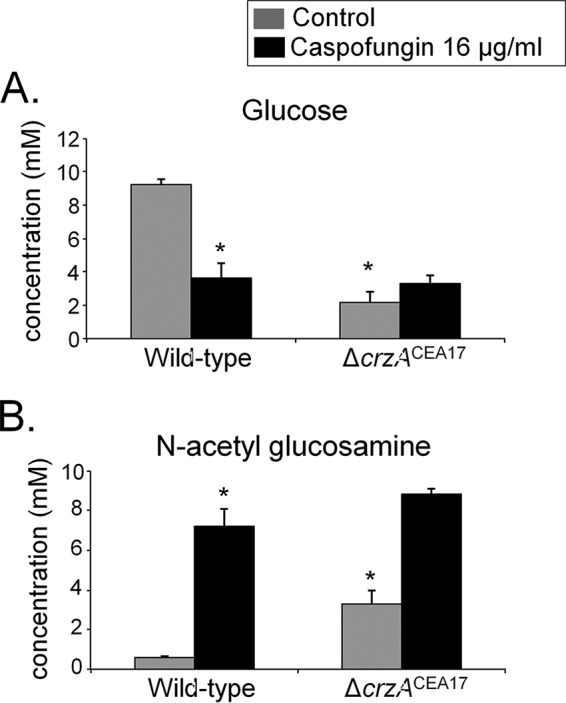
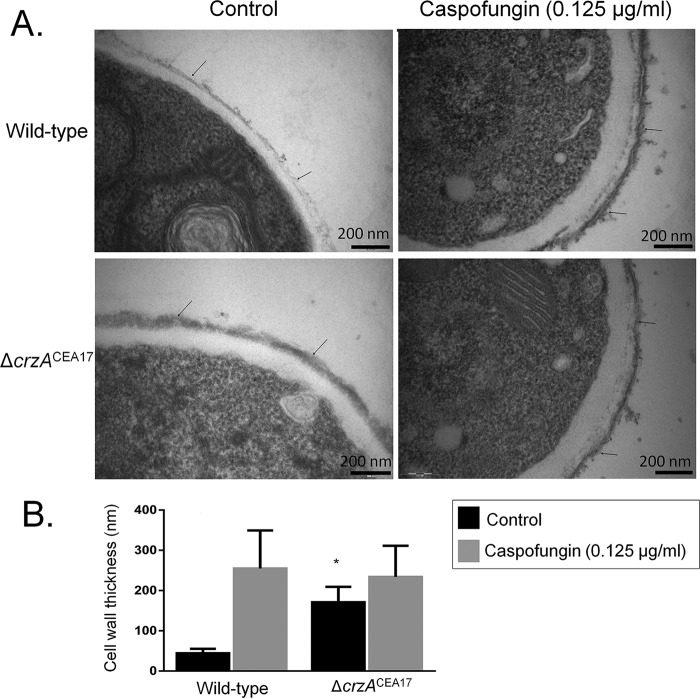
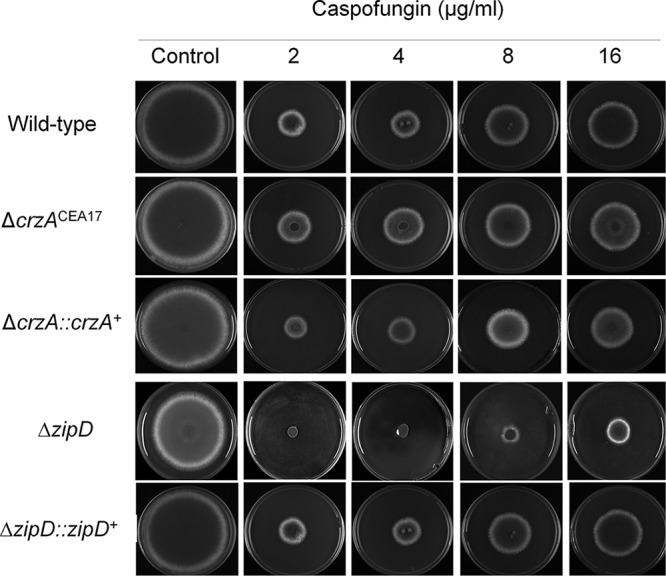
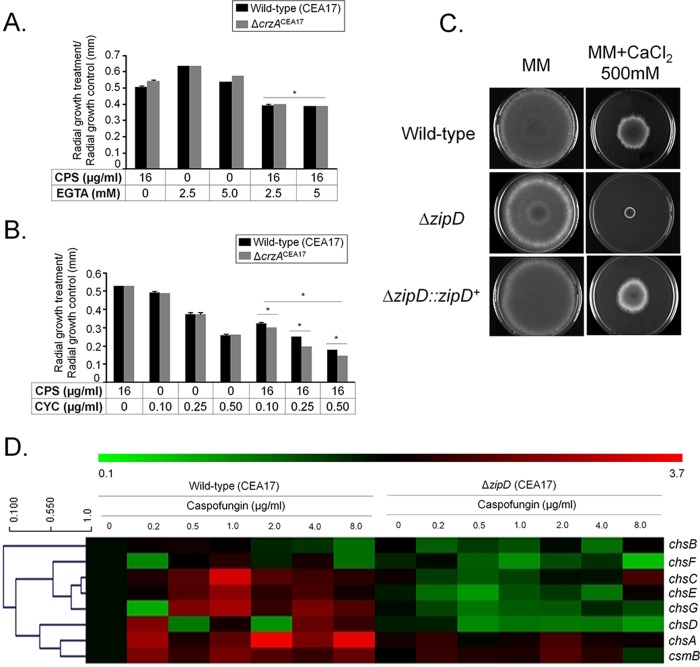
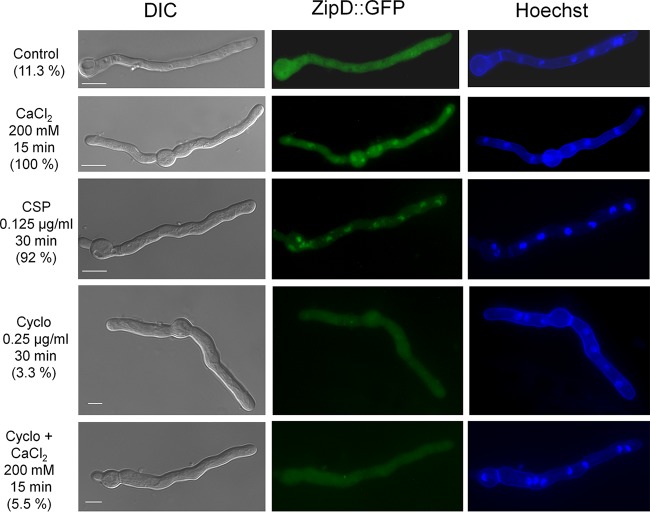
Aspergillus fumigatus is an opportunistic fungal pathogen that causes invasive aspergillosis (IA), a life-threatening disease in immunocompromised humans. The echinocandin caspofungin, adopted as a second-line therapy in combating IA, is a β-1,3-glucan synthase inhibitor, which, when used in high concentrations, reverts the anticipated A. fumigatus growth inhibition, a phenomenon called the "caspofungin paradoxical effect" (CPE). The CPE has been widely associated with increased chitin content in the cell wall due to a compensatory upregulation of chitin synthase-encoding genes. Here, we demonstrate that the CPE is dependent on the cell wall integrity (CWI) mitogen-activated protein kinase MpkAMPK1 and its associated transcription factor (TF) RlmARLM1, which regulate chitin synthase gene expression in response to different concentrations of caspofungin. Furthermore, the calcium- and calcineurin-dependent TF CrzA binds to and regulates the expression of specific chitin synthase genes during the CPE. These results suggest that the regulation of cell wall biosynthetic genes occurs by several cellular signaling pathways. In addition, CrzA is also involved in cell wall organization in the absence of caspofungin. Differences in the CPE were also observed between two A. fumigatus clinical isolates, which led to the identification of a novel basic leucine zipper TF, termed ZipD. This TF functions in the calcium-calcineurin pathway and is involved in the regulation of cell wall biosynthesis genes. This study therefore unraveled additional mechanisms and novel factors governing the CPE response, which ultimately could aid in developing more effective antifungal therapies.IMPORTANCE Systemic Aspergillus fumigatus infections are often accompanied by high mortality rates. The fungal cell wall is important for infection as it has immunomodulatory and immunoevasive properties. Paradoxical growth of A. fumigatus in the presence of high concentrations of the cell wall-disturbing agent caspofungin has been observed for more than a decade, although the mechanistic nature of this phenomenon remains largely uncharacterized. Here, we show that the CWI pathway components MpkA and RlmA as well as the calcium/calcineurin-responsive transcription factor CrzA regulate the expression of cell wall biosynthetic genes during the caspofungin paradoxical effect (CPE). Furthermore, an additional, novel calcium/calcineurin-responsive transcription factor was identified to play a role in cell wall biosynthesis gene expression during the CPE. This work paints a crucial role for calcium metabolism in the CPE and provides further insight into the complex regulation of cell wall biosynthesis, which could ultimately lead to the development of more efficient antifungal therapies.
Keywords: Aspergillus fumigatus; caspofungin; cell wall integrity pathway; paradoxical effect.
Copyright © 2017 Ries et al.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
