

Human fetal dendritic cells promote prenatal T-cell immune suppression through arginase-2
- PMID: 28614294
- PMCID: PMC6588541
- DOI: 10.1038/nature22795
Human fetal dendritic cells promote prenatal T-cell immune suppression through arginase-2
Abstract
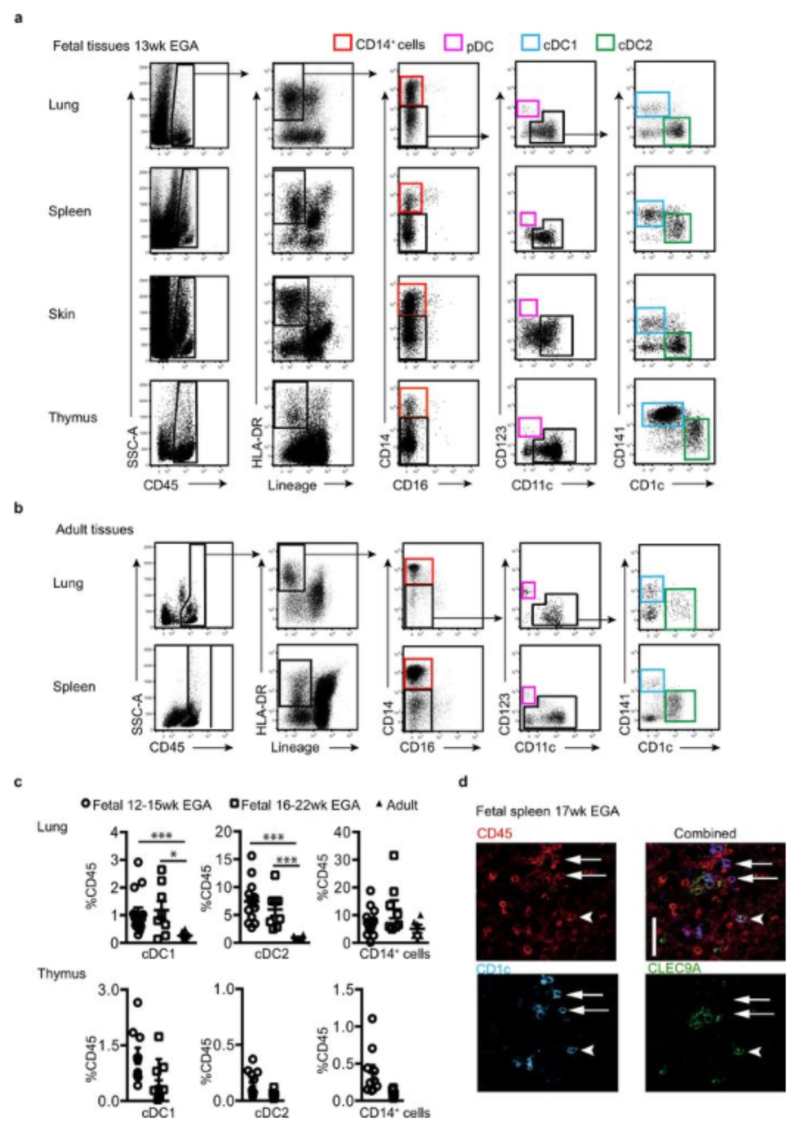
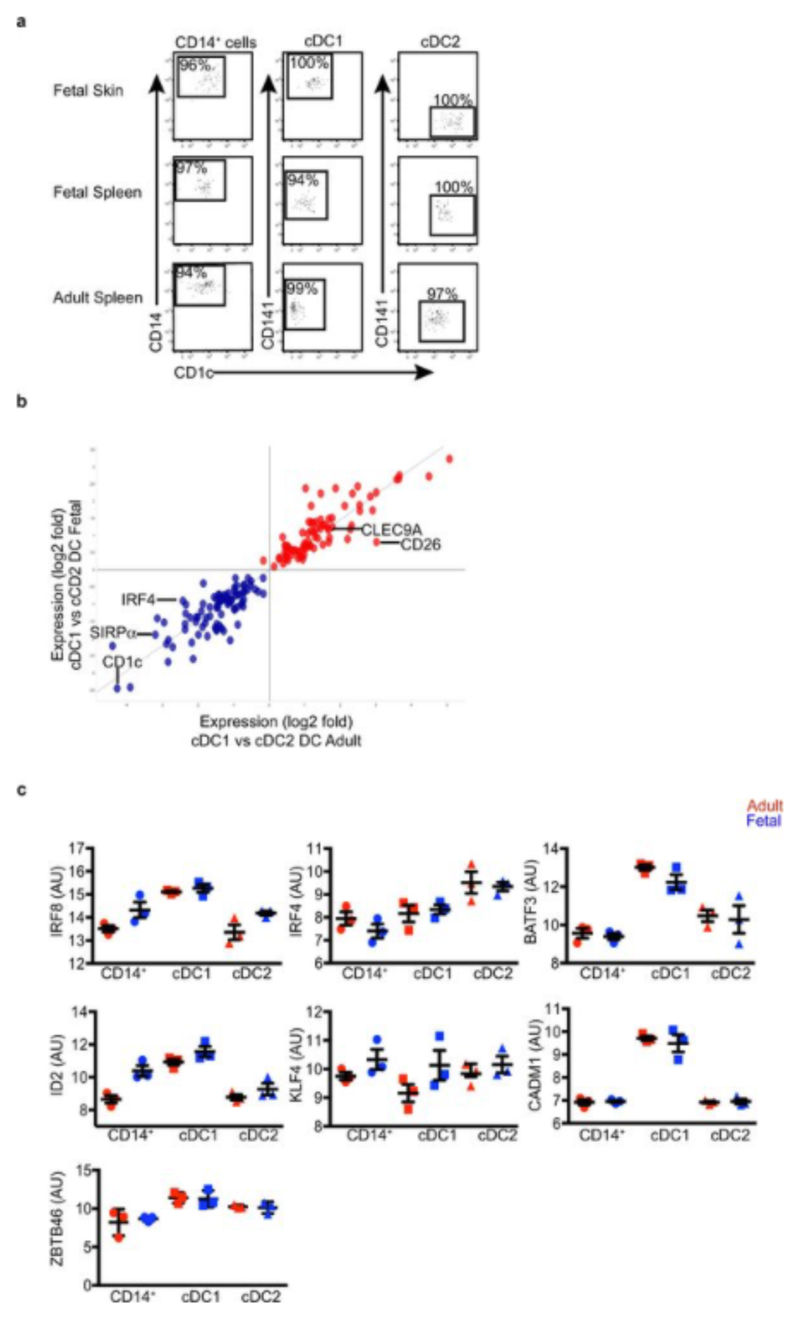
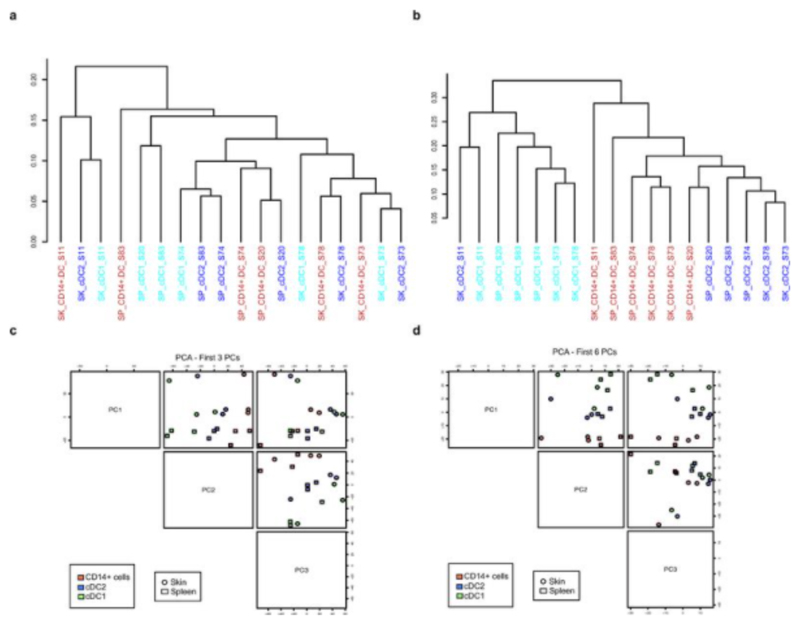
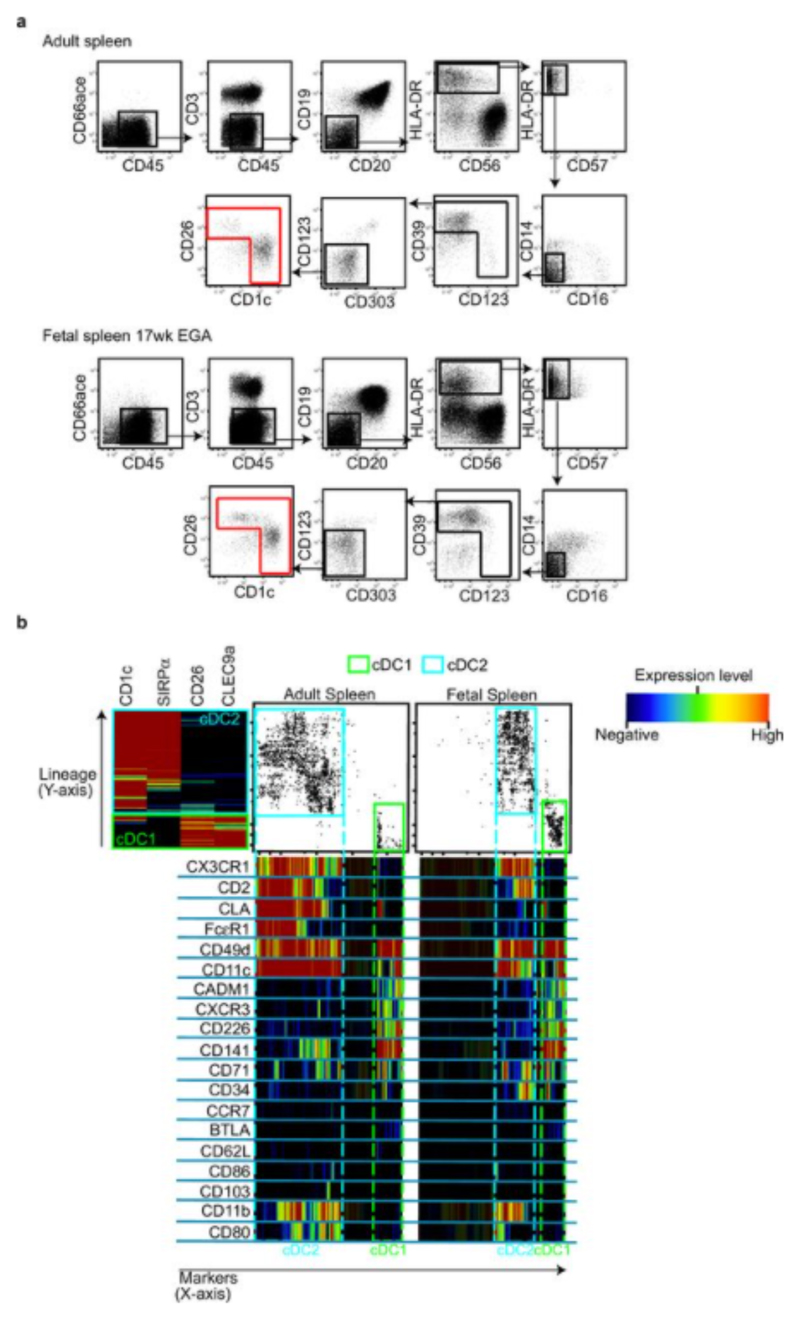
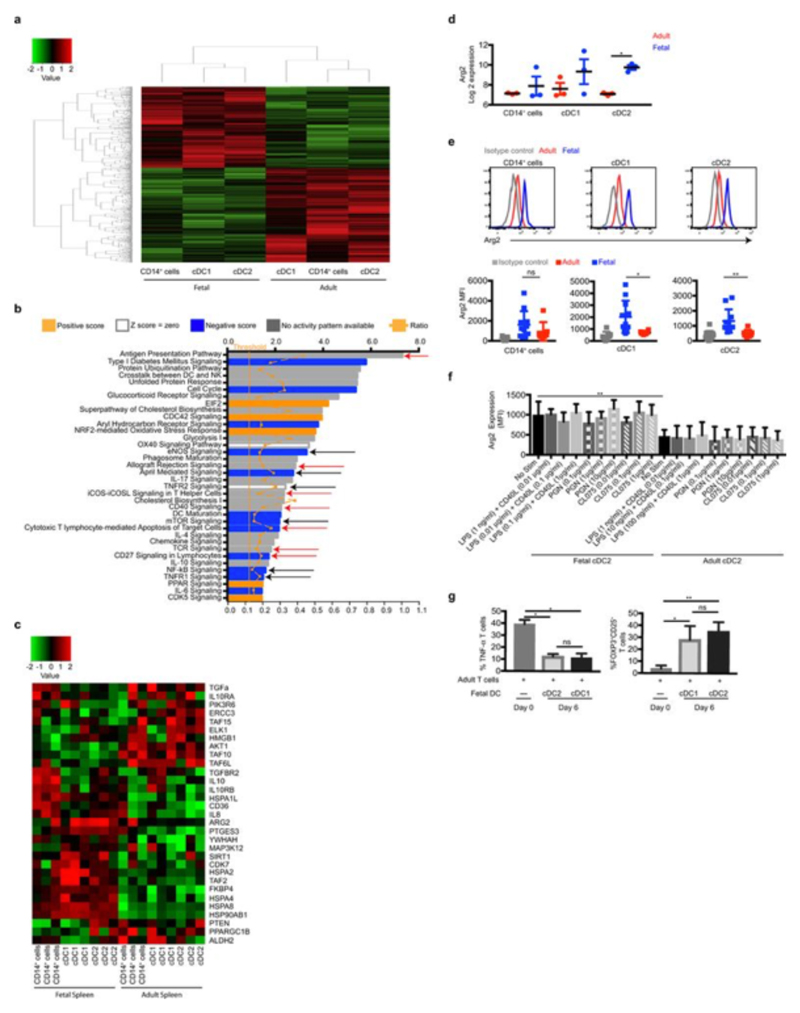
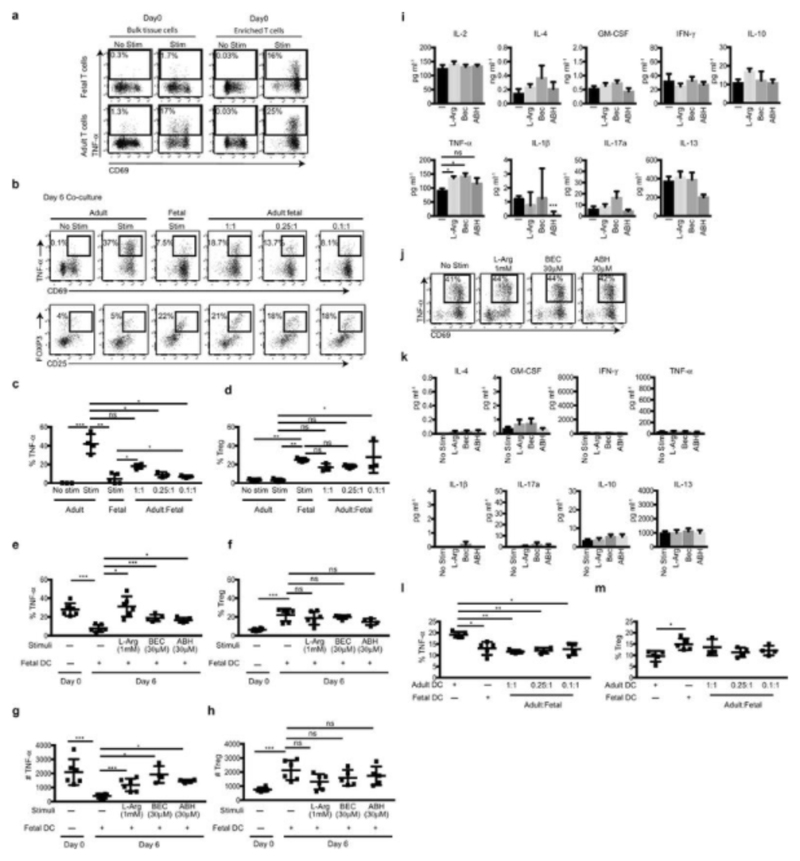
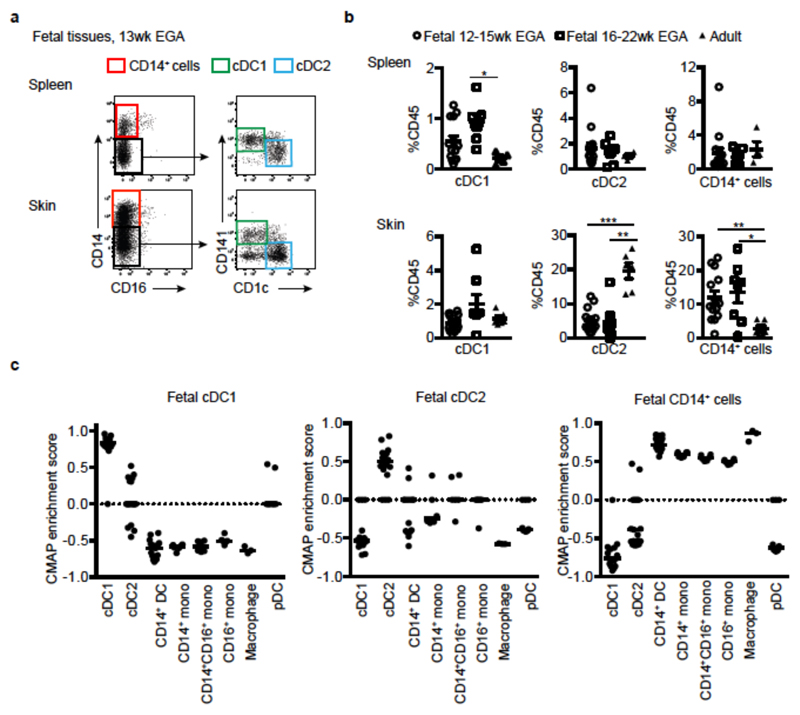
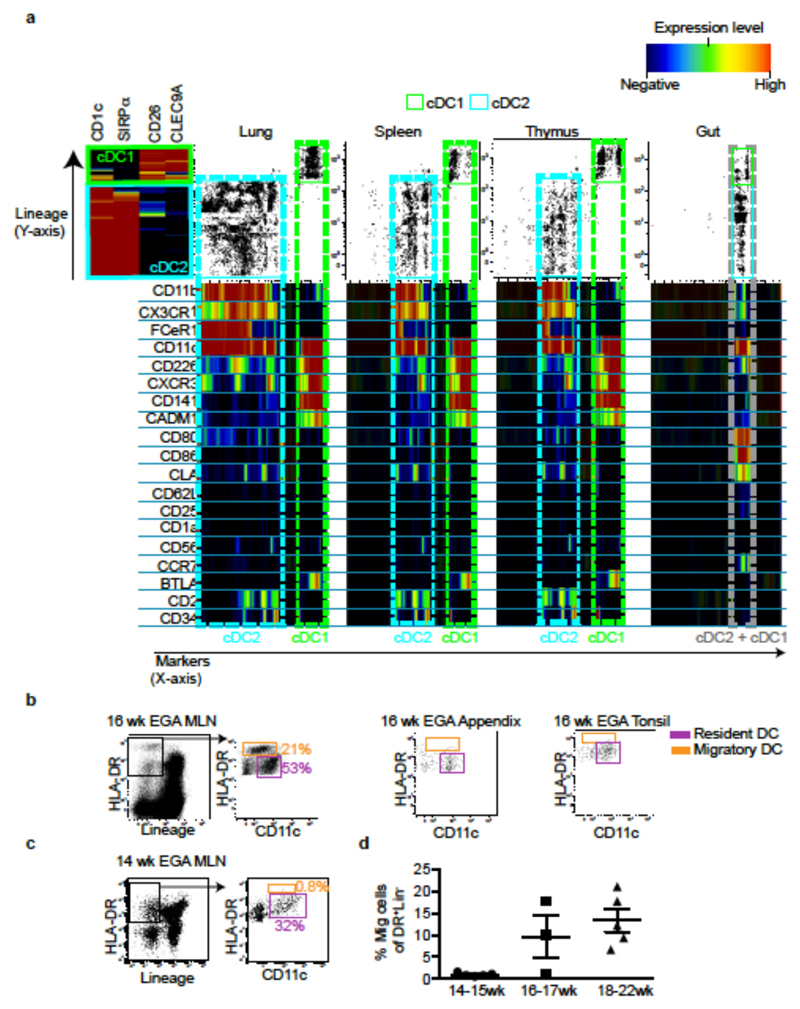
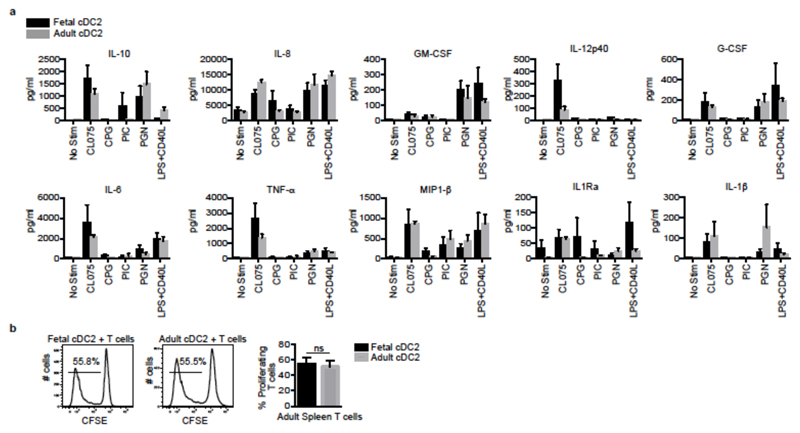
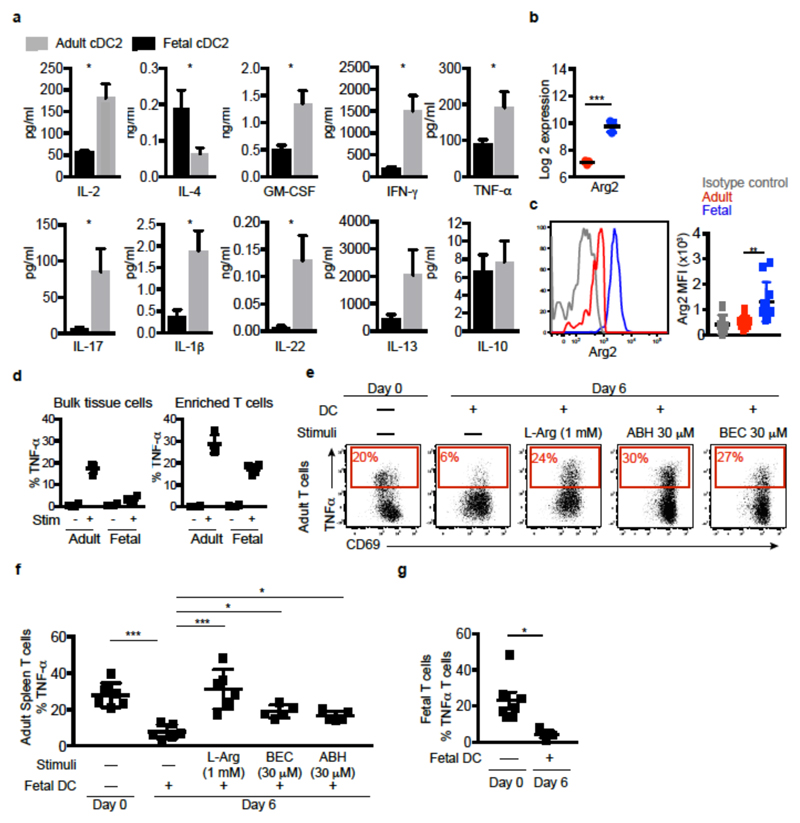
During gestation the developing human fetus is exposed to a diverse range of potentially immune-stimulatory molecules including semi-allogeneic antigens from maternal cells, substances from ingested amniotic fluid, food antigens, and microbes. Yet the capacity of the fetal immune system, including antigen-presenting cells, to detect and respond to such stimuli remains unclear. In particular, dendritic cells, which are crucial for effective immunity and tolerance, remain poorly characterized in the developing fetus. Here we show that subsets of antigen-presenting cells can be identified in fetal tissues and are related to adult populations of antigen-presenting cells. Similar to adult dendritic cells, fetal dendritic cells migrate to lymph nodes and respond to toll-like receptor ligation; however, they differ markedly in their response to allogeneic antigens, strongly promoting regulatory T-cell induction and inhibiting T-cell tumour-necrosis factor-α production through arginase-2 activity. Our results reveal a previously unappreciated role of dendritic cells within the developing fetus and indicate that they mediate homeostatic immune-suppressive responses during gestation.
Figures
Comment in
-
Eye-opening picture of fetal immune system emerges.Nature. 2017 Jun 14;546(7658):335-336. doi: 10.1038/546335a. Nature. 2017. PMID: 28617472 No abstract available.
-
Early life immunology: Fetal DCs - born to be mild.Nat Rev Immunol. 2017 Aug;17(8):465. doi: 10.1038/nri.2017.79. Epub 2017 Jul 10. Nat Rev Immunol. 2017. PMID: 28690316 No abstract available.
-
Fetal dendritic cells give mum a break.Immunol Cell Biol. 2017 Aug;95(7):575-576. doi: 10.1038/icb.2017.46. Epub 2017 Jul 18. Immunol Cell Biol. 2017. PMID: 28741621 No abstract available.
-
Formidable Defenses In Utero.Cell. 2017 Aug 10;170(4):595-597. doi: 10.1016/j.cell.2017.07.048. Cell. 2017. PMID: 28802033
References
-
- Claas FH, Gijbels Y, van der Velden-de Munck J, van Rood JJ. Induction of B cell unresponsiveness to noninherited maternal HLA antigens during fetal life. Science. 1988;241:1815–1817. - PubMed
-
- de Vries JI, Visser GH, Prechtl HF. The emergence of fetal behaviour. II. Quantitative aspects. Early Hum Dev. 1985;12:99–120. - PubMed
-
- Campbell DE, Boyle RJ, Thornton CA, Prescott SL. Mechanisms of allergic disease - environmental and genetic determinants for the development of allergy. Clin Exp Allergy. 2015;45:844–858. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
