

ERF mutations reveal a balance of ETS factors controlling prostate oncogenesis
- PMID: 28614298
- PMCID: PMC5576182
- DOI: 10.1038/nature22820
ERF mutations reveal a balance of ETS factors controlling prostate oncogenesis
Abstract
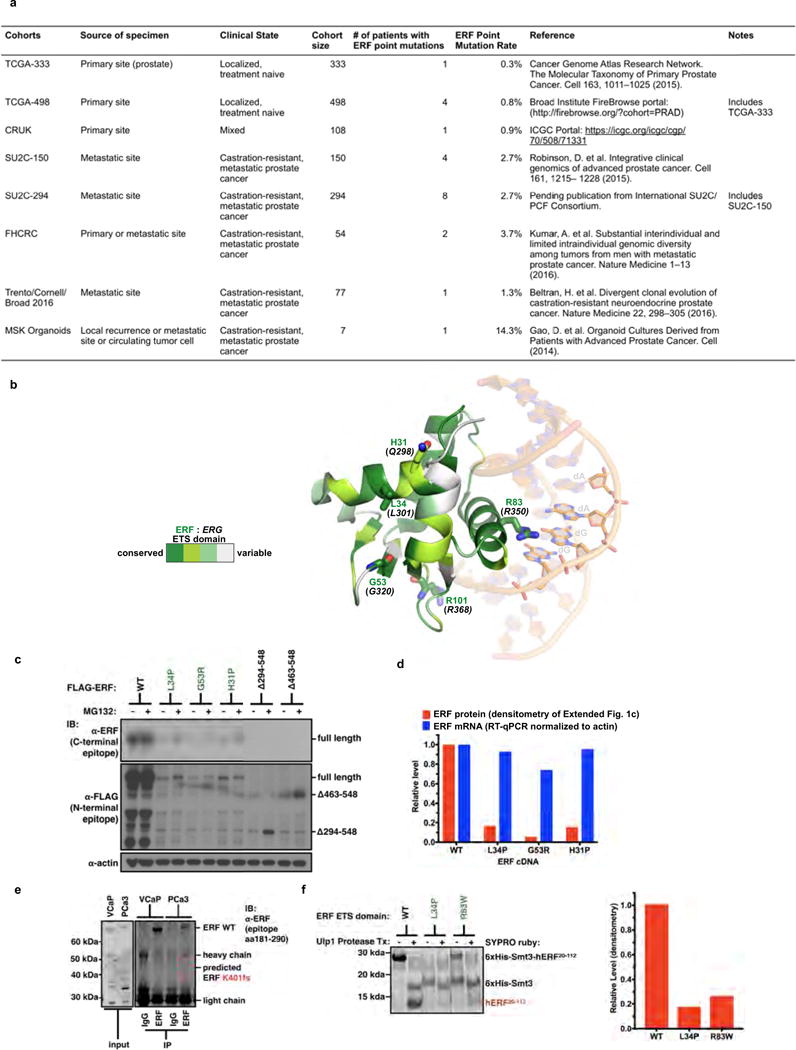
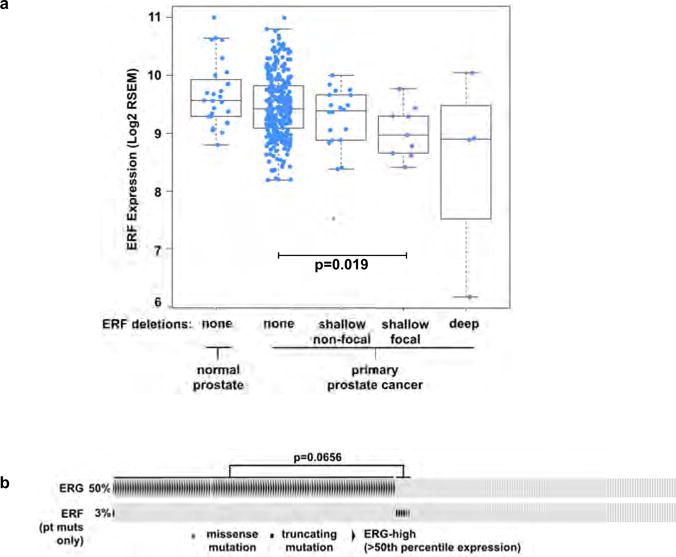
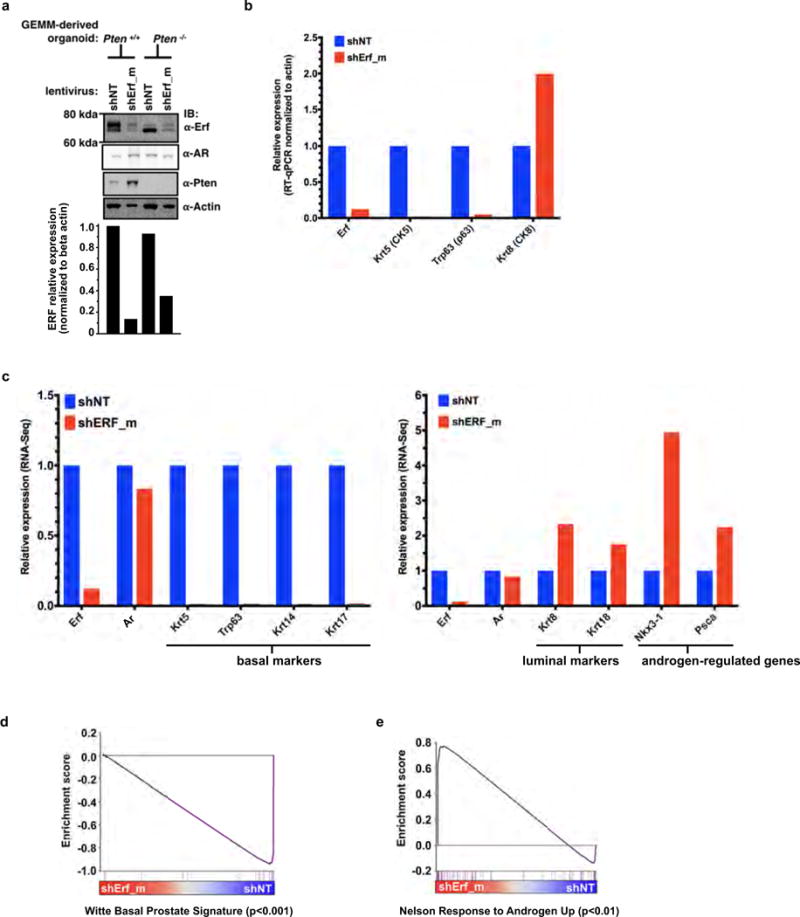
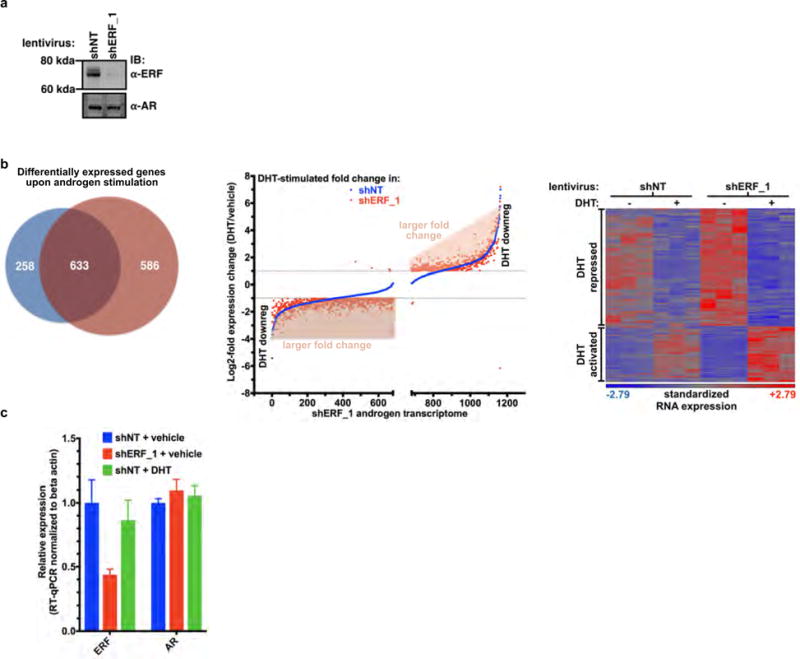
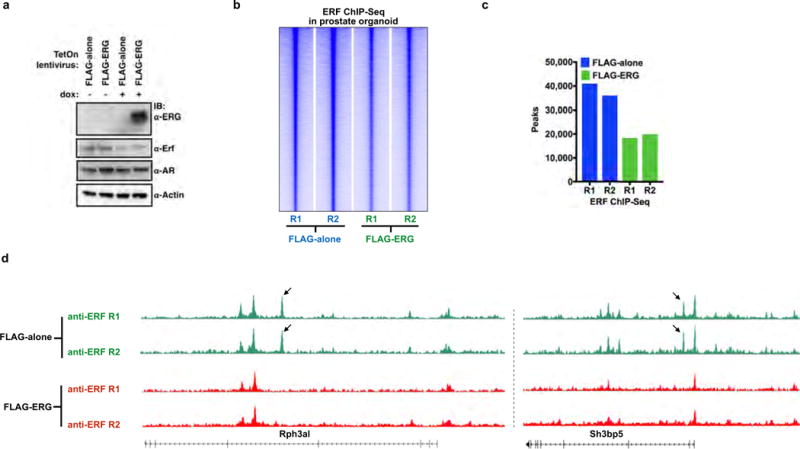
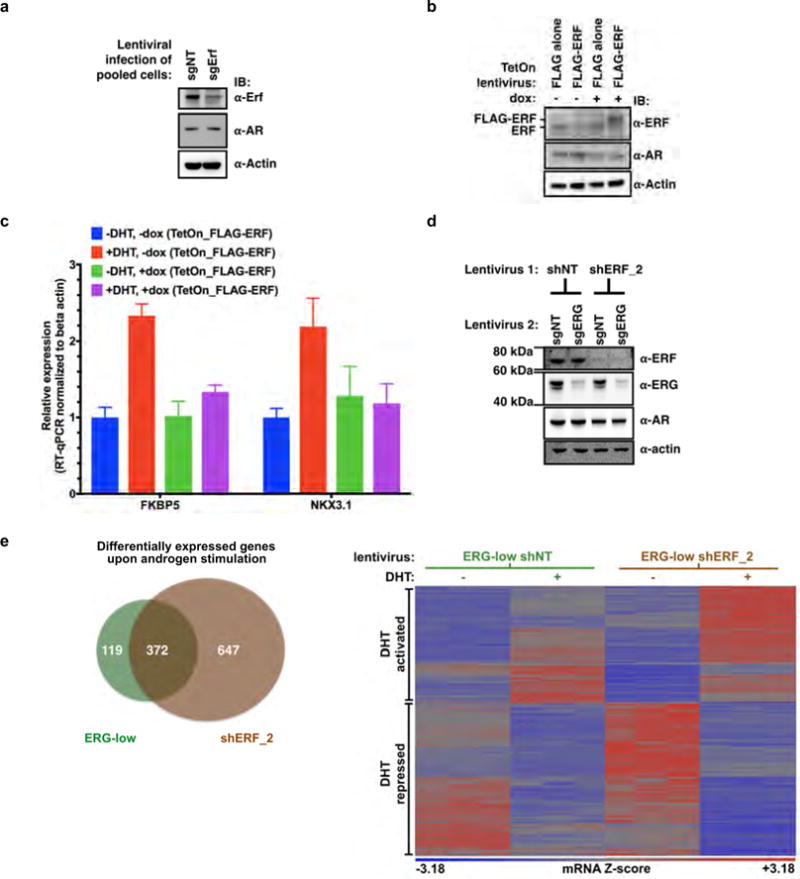
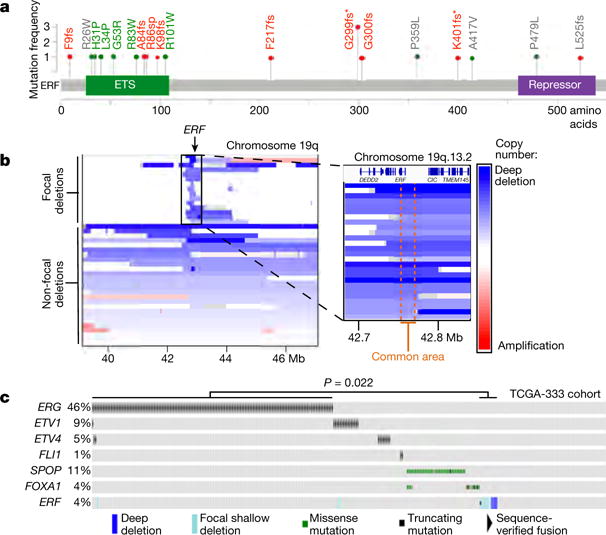
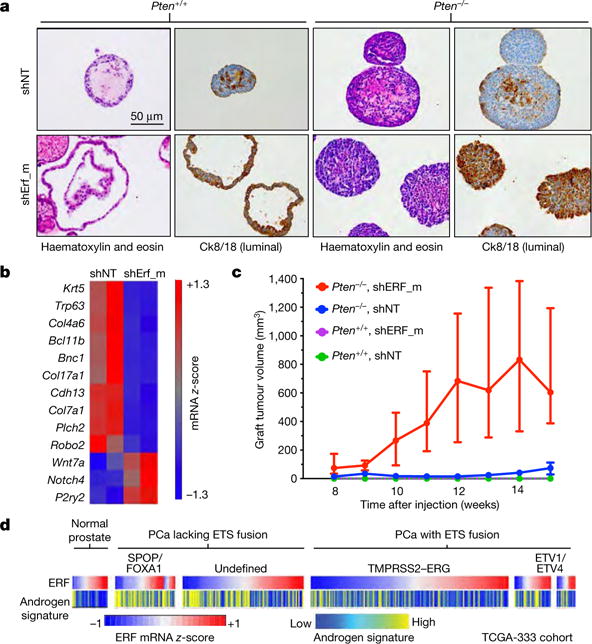
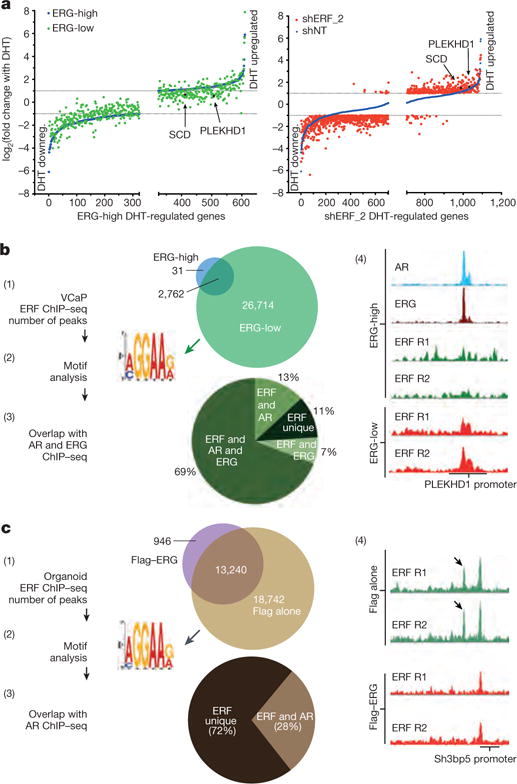
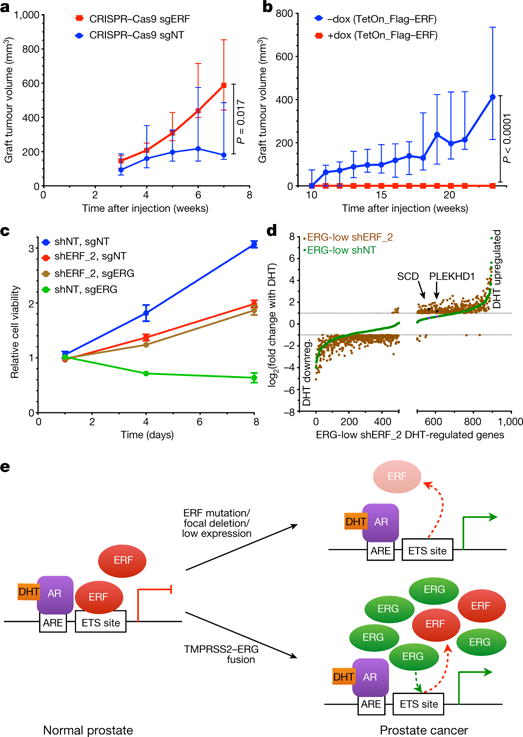
Half of all prostate cancers are caused by the TMPRSS2-ERG gene-fusion, which enables androgens to drive expression of the normally silent E26 transformation-specific (ETS) transcription factor ERG in prostate cells. Recent genomic landscape studies of such cancers have reported recurrent point mutations and focal deletions of another ETS member, the ETS2 repressor factor ERF. Here we show these ERF mutations cause decreased protein stability and mostly occur in tumours without ERG upregulation. ERF loss recapitulates the morphological and phenotypic features of ERG gain in normal mouse prostate cells, including expansion of the androgen receptor transcriptional repertoire, and ERF has tumour suppressor activity in the same genetic background of Pten loss that yields oncogenic activity by ERG. In the more common scenario of ERG upregulation, chromatin immunoprecipitation followed by sequencing indicates that ERG inhibits the ability of ERF to bind DNA at consensus ETS sites both in normal and in cancerous prostate cells. Consistent with a competition model, ERF overexpression blocks ERG-dependent tumour growth, and ERF loss rescues TMPRSS2-ERG-positive prostate cancer cells from ERG dependency. Collectively, these data provide evidence that the oncogenicity of ERG is mediated, in part, by competition with ERF and they raise the larger question of whether other gain-of-function oncogenic transcription factors might also inactivate endogenous tumour suppressors.
Conflict of interest statement
The authors declare no competing financial interests. Readers are welcome to comment on the online version of the paper. Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Comment in
-
Prostate cancer: ETS factors in the balance.Nat Rev Urol. 2017 Aug;14(8):454. doi: 10.1038/nrurol.2017.103. Epub 2017 Jun 27. Nat Rev Urol. 2017. PMID: 28654090 No abstract available.
References
-
- Tomlins SA, et al. Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer. Science. 2005;310:644–648. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
