

Stem Cells in Repair of Gastrointestinal Epithelia
- PMID: 28615312
- PMCID: PMC5545610
- DOI: 10.1152/physiol.00005.2017
Stem Cells in Repair of Gastrointestinal Epithelia
Abstract
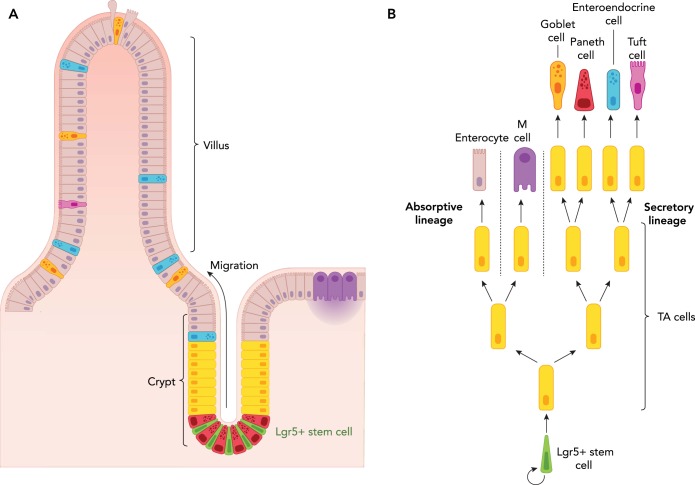
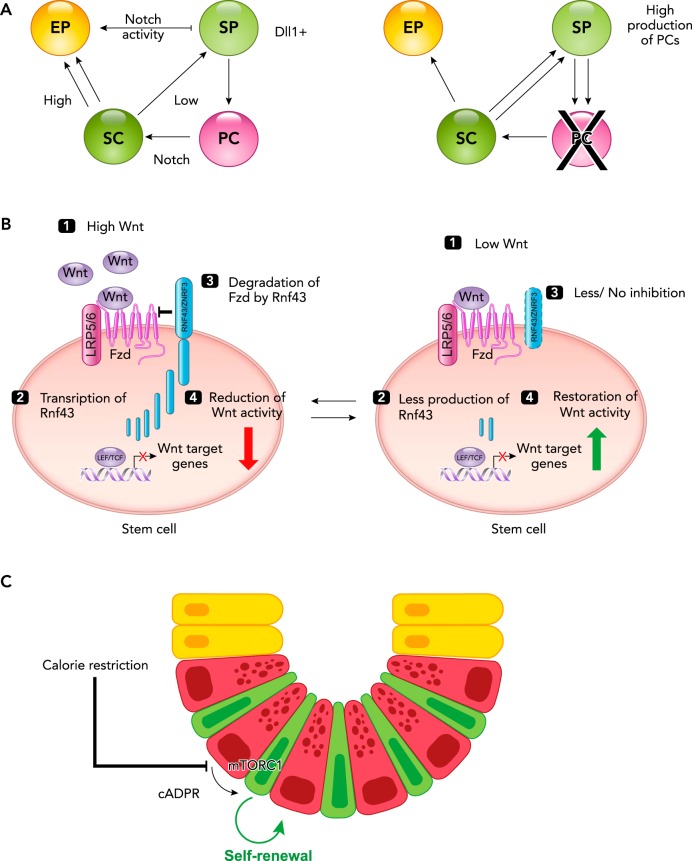
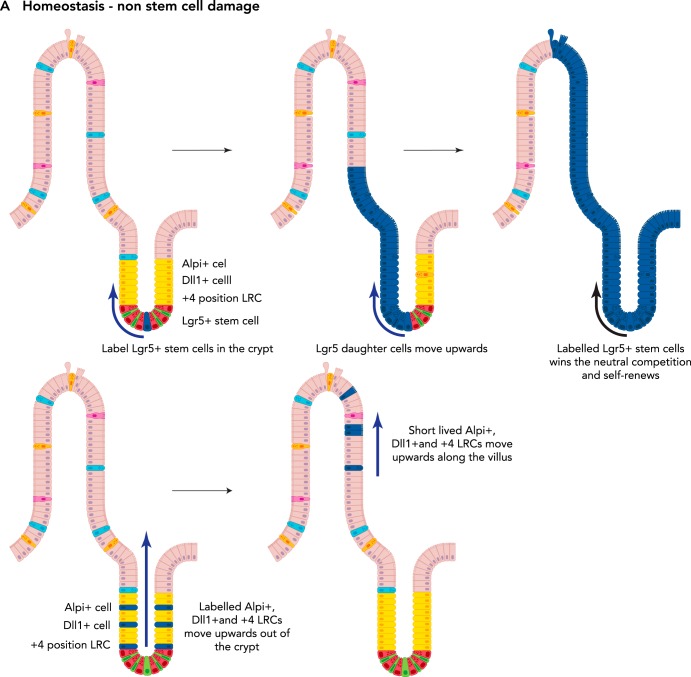
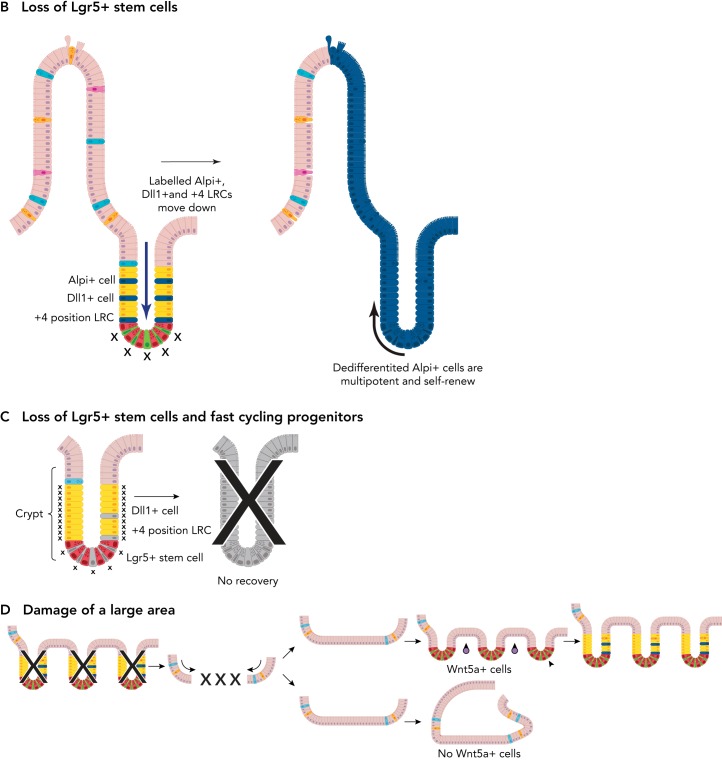
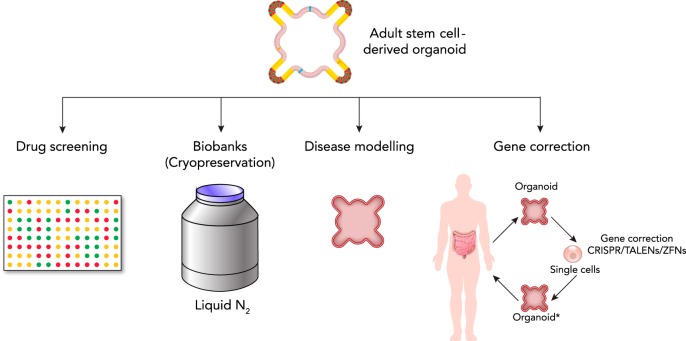
Among the endodermal tissues of adult mammals, the gastrointestinal (GI) epithelium exhibits the highest turnover rate. As the ingested food moves along the GI tract, gastric acid, digestive enzymes, and gut resident microbes aid digestion as well as nutrient and mineral absorption. Due to the harsh luminal environment, replenishment of new epithelial cells is essential to maintain organ structure and function during routine turnover and injury repair. Tissue-specific adult stem cells in the GI tract serve as a continuous source for this immense regenerative activity. Tissue homeostasis is achieved by a delicate balance between gain and loss of cells. In homeostasis, temporal tissue damage is rapidly restored by well-balanced tissue regeneration, whereas prolonged imbalance may result in diverse pathologies of homeostasis and injury repair. Starting with a summary of the current knowledge of GI tract homeostasis, we continue with providing models of acute injury and chronic diseases. Finally, we will discuss how primary organoid cultures allow new insights into the mechanisms of homeostasis, injury repair, and disease, and how this novel 3D culture system has the potential to translate into the clinic.
Copyright © 2017 Int. Union Physiol. Sci./Am. Physiol. Soc.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the author(s).
Figures
References
-
- Adolph TE, Tomczak MF, Niederreiter L, Ko H-J, Böck J, Martinez-Naves E, Glickman JN, Tschurtschenthaler M, Hartwig J, Hosomi S, Flak MB, Cusick JL, Kohno K, Iwawaki T, Billmann-Born S, Raine T, Bharti R, Lucius R, Kweon M-N, Marciniak SJ, Choi A, Hagen SJ, Schreiber S, Rosenstiel P, Kaser A, Blumberg RS. Paneth cells as a site of origin for intestinal inflammation. Nature 503: 272–276, 2013. doi: 10.1038/nature12599. - DOI - PMC - PubMed
-
- Ahn S-H, Shah YM, Inoue J, Morimura K, Kim I, Yim S, Lambert G, Kurotani R, Nagashima K, Gonzalez FJ, Inoue Y. Hepatocyte nuclear factor 4alpha in the intestinal epithelial cells protects against inflammatory bowel disease. Inflamm Bowel Dis 14: 908–920, 2008. doi: 10.1002/ibd.20413. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials