

Biogenesis and functions of mammalian iron-sulfur proteins in the regulation of iron homeostasis and pivotal metabolic pathways
- PMID: 28615439
- PMCID: PMC5546015
- DOI: 10.1074/jbc.R117.789537
Biogenesis and functions of mammalian iron-sulfur proteins in the regulation of iron homeostasis and pivotal metabolic pathways
Abstract
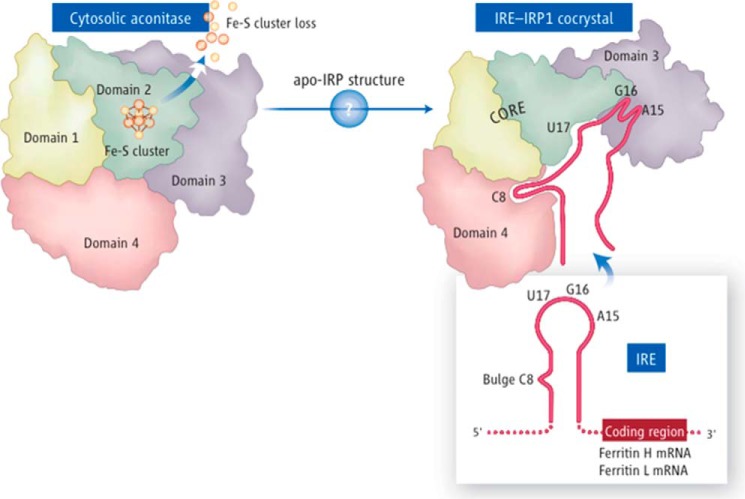
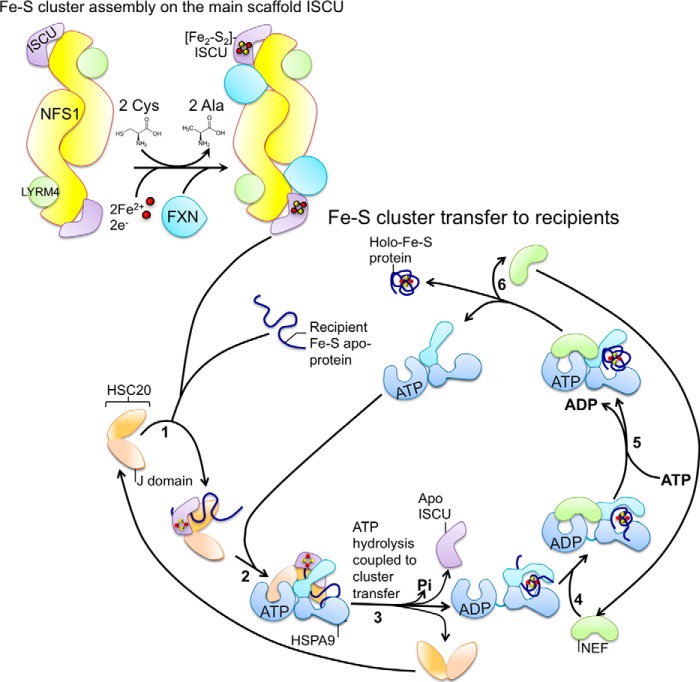
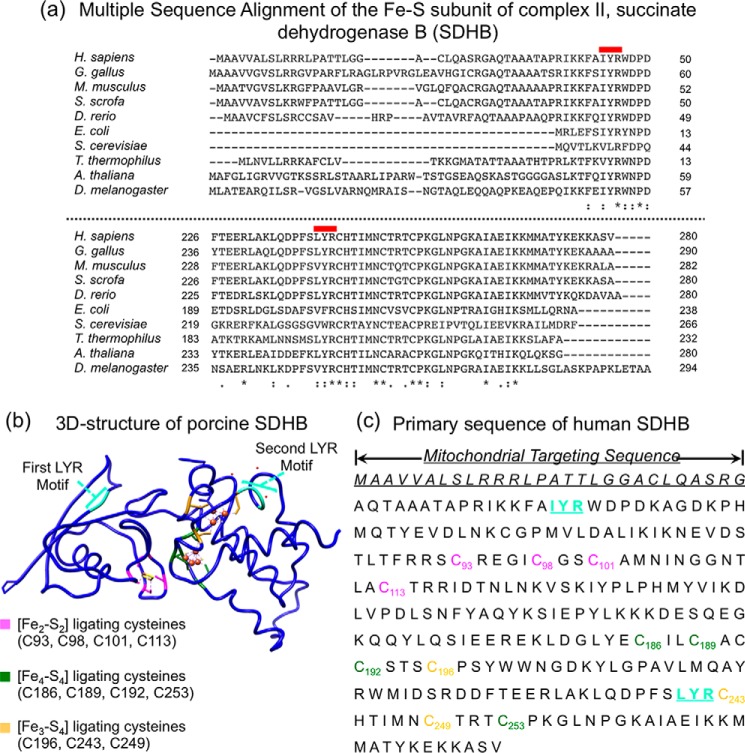
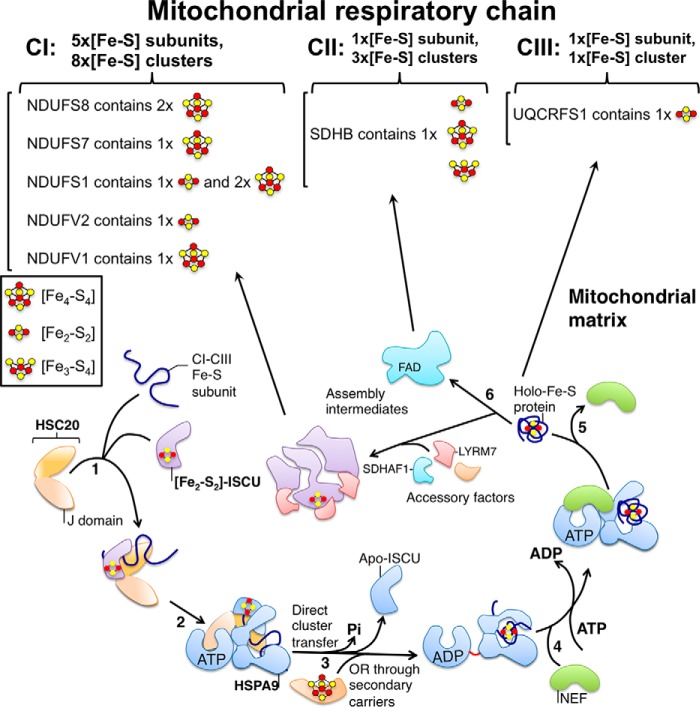
Fe-S cofactors are composed of iron and inorganic sulfur in various stoichiometries. A complex assembly pathway conducts their initial synthesis and subsequent binding to recipient proteins. In this minireview, we discuss how discovery of the role of the mammalian cytosolic aconitase, known as iron regulatory protein 1 (IRP1), led to the characterization of the function of its Fe-S cluster in sensing and regulating cellular iron homeostasis. Moreover, we present an overview of recent studies that have provided insights into the mechanism of Fe-S cluster transfer to recipient Fe-S proteins.
Keywords: HSC20; HSPA9; ISCU; energy metabolism; iron-response element (IRE); iron-sulfur cluster biogenesis; iron-sulfur protein; metalloenzyme; mitochondrial respiratory chain complex.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Zahringer J., Baliga B. S., and Munro H. N. (1979) Relative abundance of specific messenger-RNA species in the free mRNP fraction of rat liver. FEBS Lett. 108, 317–320 - PubMed
-
- Leibold E. A., Aziz N., Brown A. J., and Munro H. N. (1984) Conservation in rat liver of light and heavy subunit sequences of mammalian ferritin. Presence of unique octopeptide in the light subunit. J. Biol. Chem. 259, 4327–4334 - PubMed
-
- Hentze M. W., Caughman S. W., Rouault T. A., Barriocanal J. G., Dancis A., Harford J. B., and Klausner R. D. (1987) Identification of the iron-responsive element for the translational regulation of human ferritin mRNA. Science 238, 1570–1573 - PubMed
-
- Rouault T. A., Tang C. K., Kaptain S., Burgess W. H., Haile D. J., Samaniego F., McBride O. W., Harford J. B., and Klausner R. D. (1990) Cloning of the cDNA encoding an RNA regulatory protein–the human iron-responsive element-binding protein. Proc. Natl. Acad. Sci. U.S.A. 87, 7958–7962 - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
