

The CCAAT-Binding Complex Controls Respiratory Gene Expression and Iron Homeostasis in Candida Glabrata
- PMID: 28615656
- PMCID: PMC5471220
- DOI: 10.1038/s41598-017-03750-5
The CCAAT-Binding Complex Controls Respiratory Gene Expression and Iron Homeostasis in Candida Glabrata
Abstract
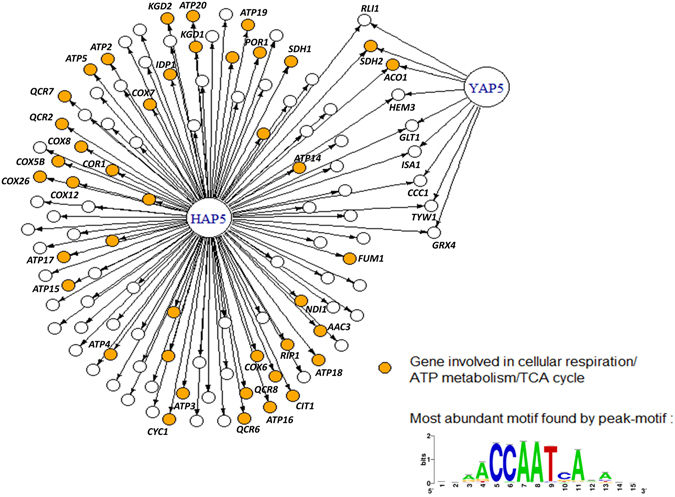
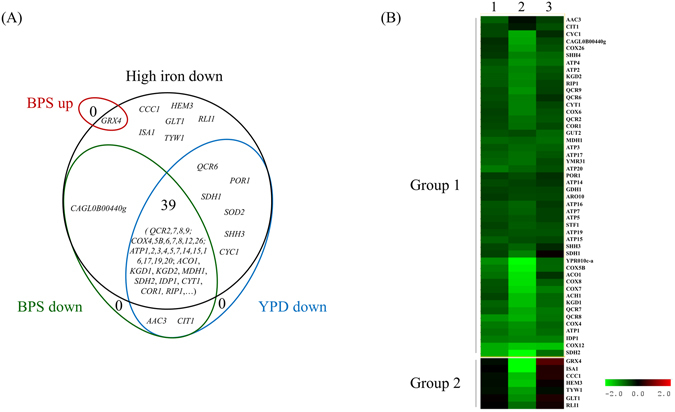
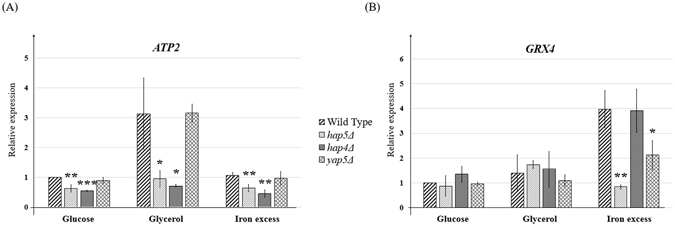
The CCAAT-binding complex (CBC) is a heterotrimeric transcription factor which is widely conserved in eukaryotes. In the model yeast S. cerevisiae, CBC positively controls the expression of respiratory pathway genes. This role involves interactions with the regulatory subunit Hap4. In many pathogenic fungi, CBC interacts with the HapX regulatory subunit to control iron homeostasis. HapX is a bZIP protein which only shares with Hap4 the Hap4Like domain (Hap4L) required for its interaction with CBC. Here, we show that CBC has a dual role in the pathogenic yeast C. glabrata. It is required, along with Hap4, for the constitutive expression of respiratory genes and it is also essential for the iron stress response, which is mediated by the Yap5 bZIP transcription factor. Interestingly, Yap5 contains a vestigial Hap4L domain. The mutagenesis of this domain severely reduced Yap5 binding to its targets and compromised its interaction with Hap5. Hence, Yap5, like HapX in other species, acts as a CBC regulatory subunit in the regulation of iron stress response. This work reveals new aspects of iron homeostasis in C. glabrata and of the evolution of the role of CBC and Hap4L-bZIP proteins in this process.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
