

Characterization of the regulation of a plant polysaccharide utilization operon and its role in biofilm formation in Bacillus subtilis
- PMID: 28617843
- PMCID: PMC5472308
- DOI: 10.1371/journal.pone.0179761
Characterization of the regulation of a plant polysaccharide utilization operon and its role in biofilm formation in Bacillus subtilis
Abstract
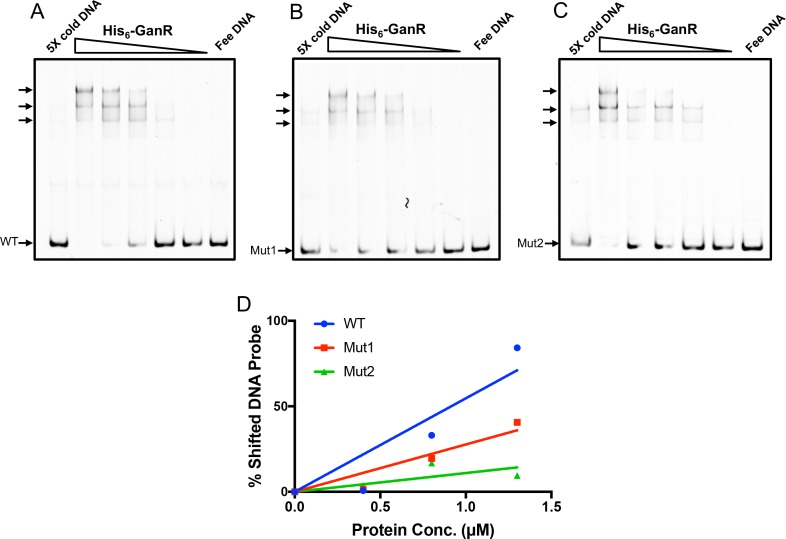
The soil bacterium Bacillus subtilis is often found in association with plants in the rhizosphere. Previously, plant polysaccharides have been shown to stimulate formation of root-associated multicellular communities, or biofilms, in this bacterium, yet the underlying mechanism is not fully understood. A five-gene gan operon (ganSPQAB) in B. subtilis has recently been shown to be involved in utilization of the plant-derived polysaccharide galactan. Despite these findings, molecular details about the regulation of the operon and the role of the operon in biofilm formation remain elusive. In this study, we performed comprehensive genetic analyses on the regulation of the gan operon. We show that this operon is regulated both by a LacI-like transcription repressor (GanR), which directly binds to pairs of inverted DNA repeats in the promoter region of the operon, and by the catabolite control protein A (CcpA). Derepression can be triggered by the presence of the inducer β-1,4-galactobiose, a hydrolysis product of galactan, or in situ when B. subtilis cells are associated with plant roots. In addition to the transcriptional regulation, the encoded ß-galactosidase GanA (by ganA), which hydrolyzes ß-1,4-galactobiose into galactose, is inhibited at the enzymatic level by the catalytic product galactose. Thus, the galactan utilization pathway is under complex regulation involving both positive and negative feedback mechanisms in B. subtilis. We discuss about the biological significance of such complex regulation as well as a hypothesis of biofilm induction by galactan via multiple mechanisms.
Conflict of interest statement
Figures





References
-
- Lugtenberg B, Kamilova F. Plant-Growth-Promoting Rhizobacteria. Annu Rev Microbiol. 2009;63(1):541–56. doi: 10.1146/annurev.micro.62.081307.162918 . - DOI - PubMed
-
- Earl AM, Losick R, Kolter R. Ecology and genomics of Bacillus subtilis. Trends in Microbiology. 2008;16(6):269–75. doi: 10.1016/j.tim.2008.03.004 - DOI - PMC - PubMed
-
- Ochiai A, Itoh T, Kawamata A, Hashimoto W, Murata K. Plant Cell Wall Degradation by Saprophytic Bacillus subtilis Strains: Gene Clusters Responsible for Rhamnogalacturonan Depolymerization. Applied and Environmental Microbiology. 2007;73(12):3803–13. doi: 10.1128/AEM.00147-07 - DOI - PMC - PubMed
-
- Sakamoto T, Hours A-R, Sakai T. Purification, Characterization, and Production of Two Pectic Transeliminases with Protopectinase Activity from Bacillus subtilis. Bioscience, Biotechnology, and Biochemistry. 1994;58(2):353–8. doi: 10.1271/bbb.58.353 - DOI - PubMed
-
- Soriano M, Diaz P, Pastor FIJ. Pectate lyase C from Bacillus subtilis: a novel endo-cleaving enzyme with activity on highly methylated pectin. Microbiology. 2006;152(3):617–25. doi: 10.1099/mic.0.28562–0 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
