

IL-22 Upregulates Epithelial Claudin-2 to Drive Diarrhea and Enteric Pathogen Clearance
- PMID: 28618266
- PMCID: PMC5541253
- DOI: 10.1016/j.chom.2017.05.009
IL-22 Upregulates Epithelial Claudin-2 to Drive Diarrhea and Enteric Pathogen Clearance
Abstract
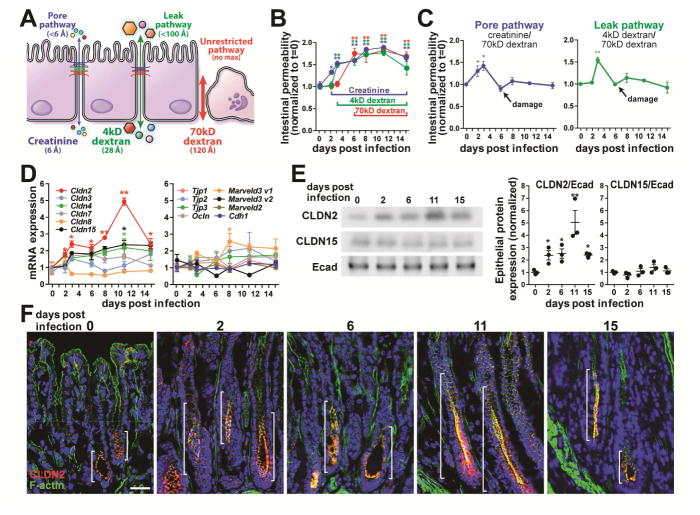
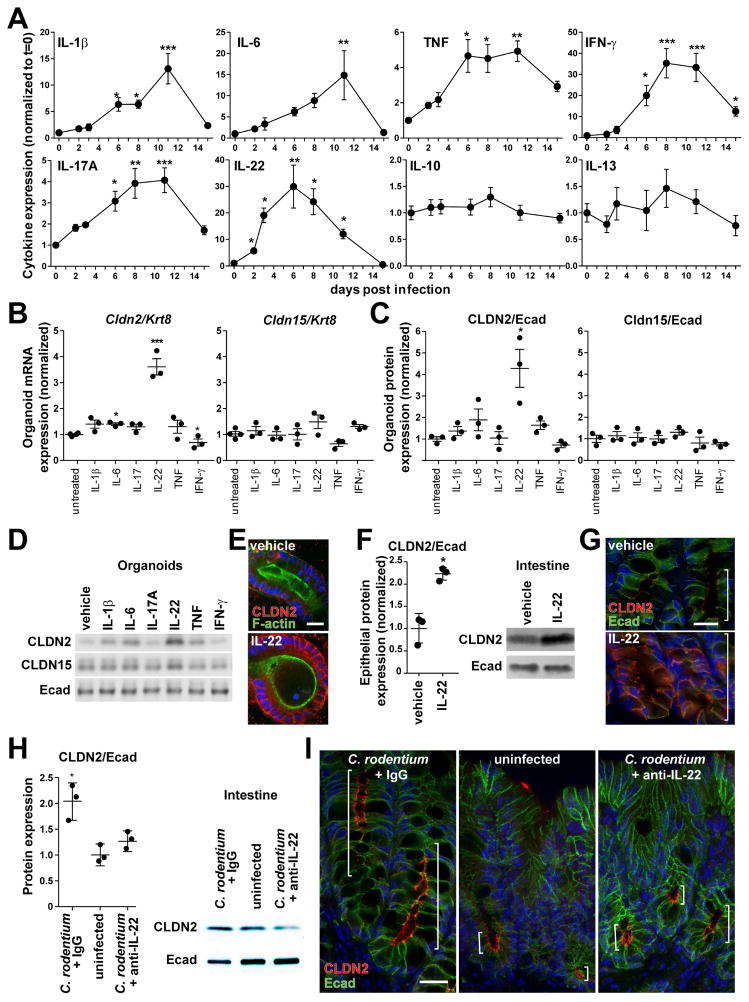
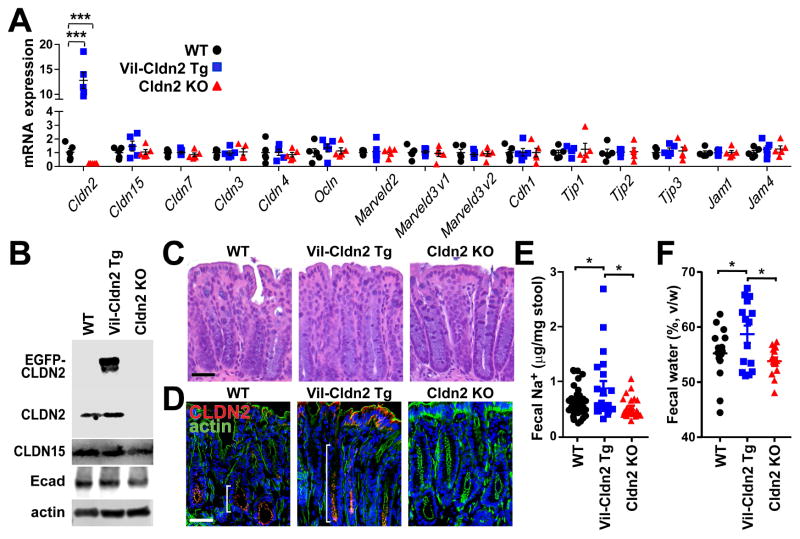
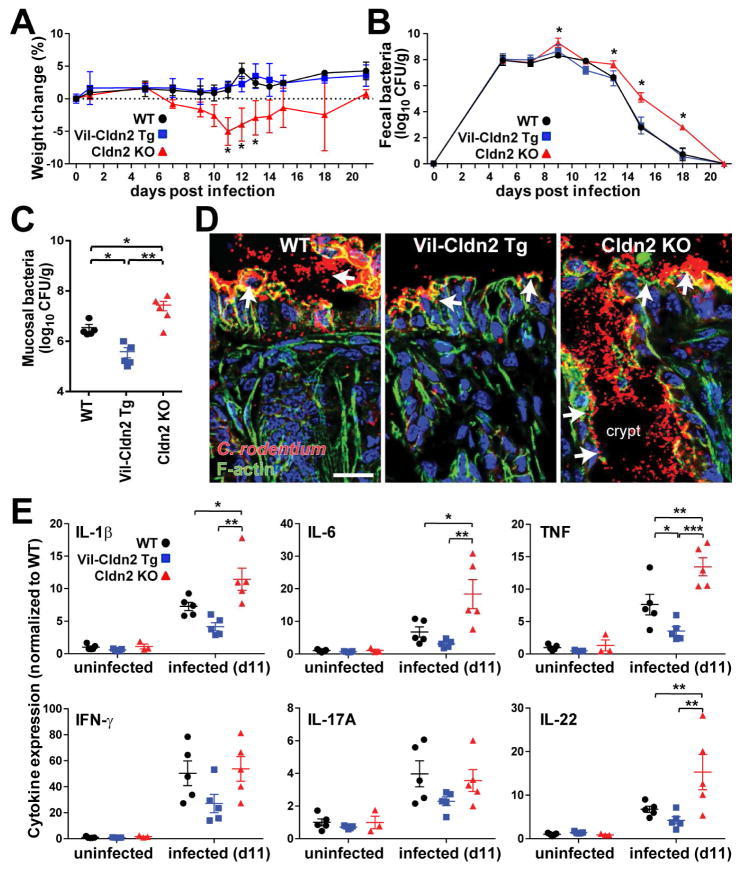
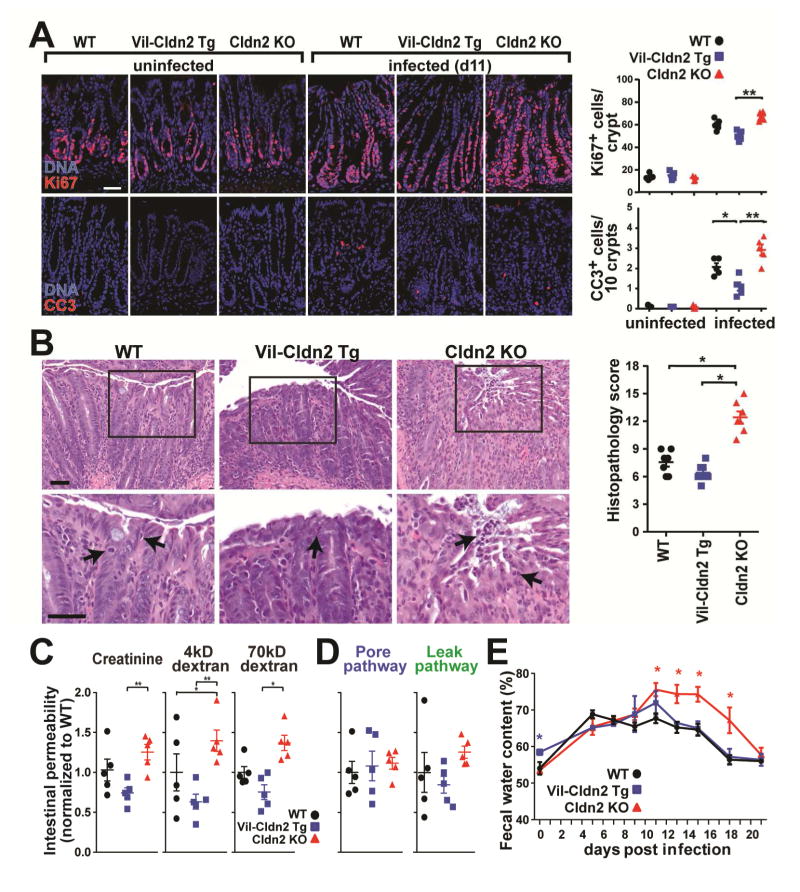
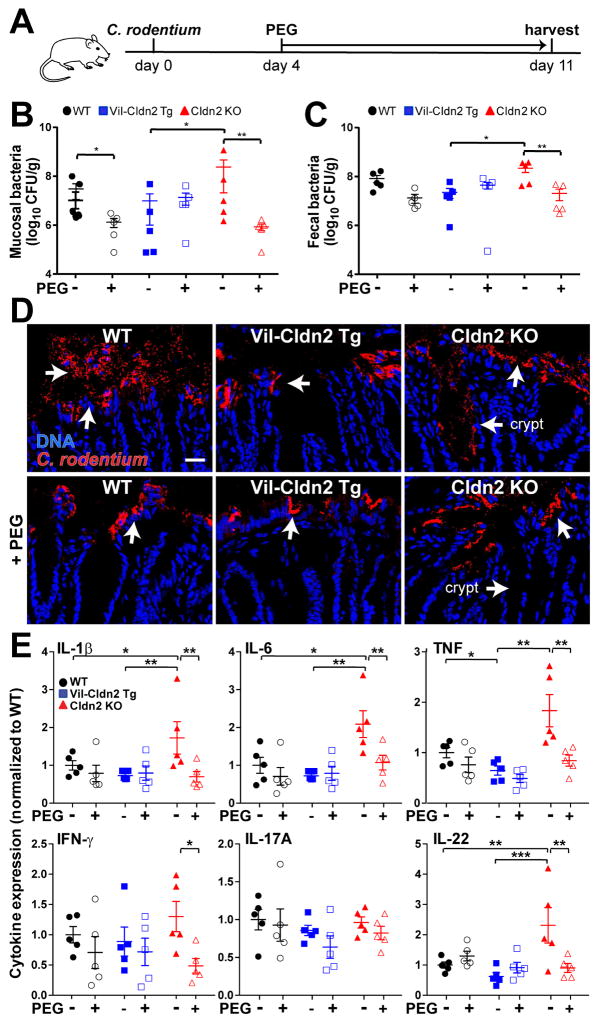
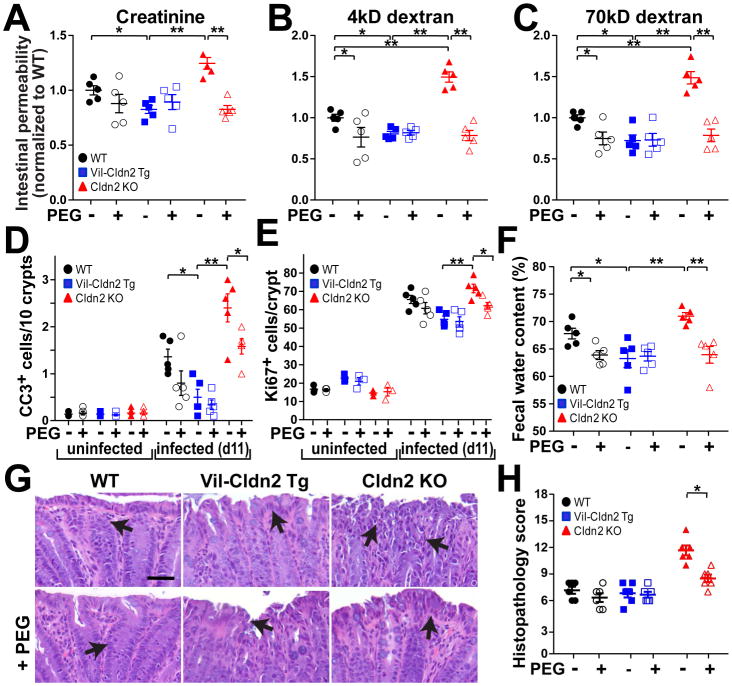
Diarrhea is a host response to enteric pathogens, but its impact on pathogenesis remains poorly defined. By infecting mice with the attaching and effacing bacteria Citrobacter rodentium, we defined the mechanisms and contributions of diarrhea and intestinal barrier loss to host defense. Increased permeability occurred within 2 days of infection and coincided with IL-22-dependent upregulation of the epithelial tight junction protein claudin-2. Permeability increases were limited to small molecules, as expected for the paracellular water and Na+ channel formed by claudin-2. Relative to wild-type, claudin-2-deficient mice experienced severe disease, including increased mucosal colonization by C. rodentium, prolonged pathogen shedding, exaggerated cytokine responses, and greater tissue injury. Conversely, transgenic claudin-2 overexpression reduced disease severity. Chemically induced osmotic diarrhea reduced colitis severity and C. rodentium burden in claudin-2-deficient, but not transgenic, mice, demonstrating that claudin-2-mediated protection is the result of enhanced water efflux. Thus, IL-22-induced claudin-2 upregulation drives diarrhea and pathogen clearance.
Keywords: bacterial infection; colitis; diarrhea; enteric infection; innate defense; permeability; tight junction.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Ahmad R, Chaturvedi R, Olivares-Villagomez D, Habib T, Asim M, Shivesh P, Polk DB, Wilson KT, Washington MK, Van Kaer L, et al. Targeted colonic claudin-2 expression renders resistance to epithelial injury, induces immune suppression, and protects from colitis. Mucosal Immunol. 2014;7:1340–1353. - PMC - PubMed
-
- Davison WC. A Bacteriological and Clinical Consideration of Bacillary Dysentery in Adults and Children. Medicine. 1922;1:389–510.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
