

Mechanisms underlying extensive Ser129-phosphorylation in α-synuclein aggregates
- PMID: 28619113
- PMCID: PMC5472914
- DOI: 10.1186/s40478-017-0452-6
Mechanisms underlying extensive Ser129-phosphorylation in α-synuclein aggregates
Abstract
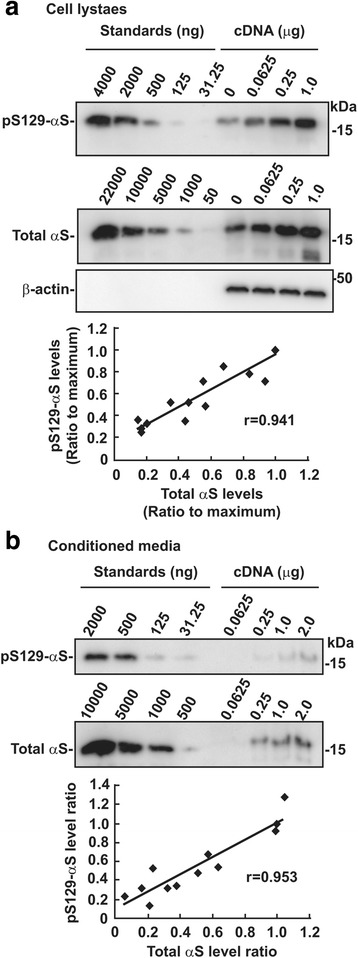
Parkinson's disease (PD) is characterized neuropathologically by intracellular aggregates of fibrillar α-synuclein, termed Lewy bodies (LBs). Approximately 90% of α-synuclein deposited as LBs is phosphorylated at Ser129 in brains with PD. In contrast, only 4% of total α-synuclein is phosphorylated at Ser129 in brains with normal individuals. It is unclear why extensive phosphorylation occurs in the pathological process of PD. To address this issue, we investigated a mechanism and role of Ser129-phosphorylation in regulating accumulation of α-synuclein. In CHO cells, the levels of Ser129-phosphorylated soluble α-synuclein were maintained constantly to those of total α-synuclein in intracellular and extracellular spaces. In SH-SY5Y cells and rat primary cortical neurons, mitochondrial impairment by rotenone or MPP+ enhanced Ser129-phosphorylation through increased influx of extracellular Ca2+. This elevation was suppressively controlled by targeting Ser129-phosphorylated α-synuclein to the proteasome pathway. Rotenone-induced insoluble α-synuclein was also targeted by Ser129-phosphoryation to the proteasome pathway. Experiments with epoxomicin and chloroquine showed that proteasomal targeting of insoluble Ser129-phosphorylated α-synuclein was enhanced under lysosome inhibition and it reduced accumulation of insoluble total α-synuclein. However, in a rat AAV-mediated α-synuclein overexpression model, there was no difference in the number of total α-synuclein aggregates between A53T mutant and A53T plus S129A double mutant α-synuclein, although Ser129-phosphorylated α-synuclein-positive aggregates were increased in rats expressing A53T α-synuclein. These findings suggest that Ser129-phosphorylation occurs against stress conditions, which increases influx of extracellular Ca2+, and it prevents accumulation of insoluble α-synuclein by evoking proteasomal clearance complementary to lysosomal one. However, Ser129-phosphorylation may provide an ineffective signal for degradation-resistant aggregates, causing extensive phosphorylation in aggregates.
Keywords: Aggregation; Mitochondrial impairment; Parkinson’s disease; Phosphorylation; Proteasome pathway; α–Synuclein.
Figures








References
-
- Anderson JP, Walker DE, Goldstein JM, de Laat R, Banducci K, Caccavello RJ, Barbour R, Huang J, Kling K, Lee M, et al. Phosphorylation of ser-129 is the dominant pathological modification of alpha-synuclein in familial and sporadic Lewy body disease. J Biol Chem. 2006;281:29739–29752. doi: 10.1074/jbc.M600933200. - DOI - PubMed
-
- Arawaka S, Fukushima S, Sato H, Sasaki A, Koga K, Koyama S, Kato T (2014) Zonisamide attenuates alpha-synuclein neurotoxicity by an aggregation-independent mechanism in a rat model of familial Parkinson’s disease. PLoS One 9:e89076. https://doi.org/10.1371/journal.pone.0089076 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
