

Using CRISPR-Cas9 to Generate Gene-Corrected Autologous iPSCs for the Treatment of Inherited Retinal Degeneration
- PMID: 28619647
- PMCID: PMC5589061
- DOI: 10.1016/j.ymthe.2017.05.015
Using CRISPR-Cas9 to Generate Gene-Corrected Autologous iPSCs for the Treatment of Inherited Retinal Degeneration
Abstract
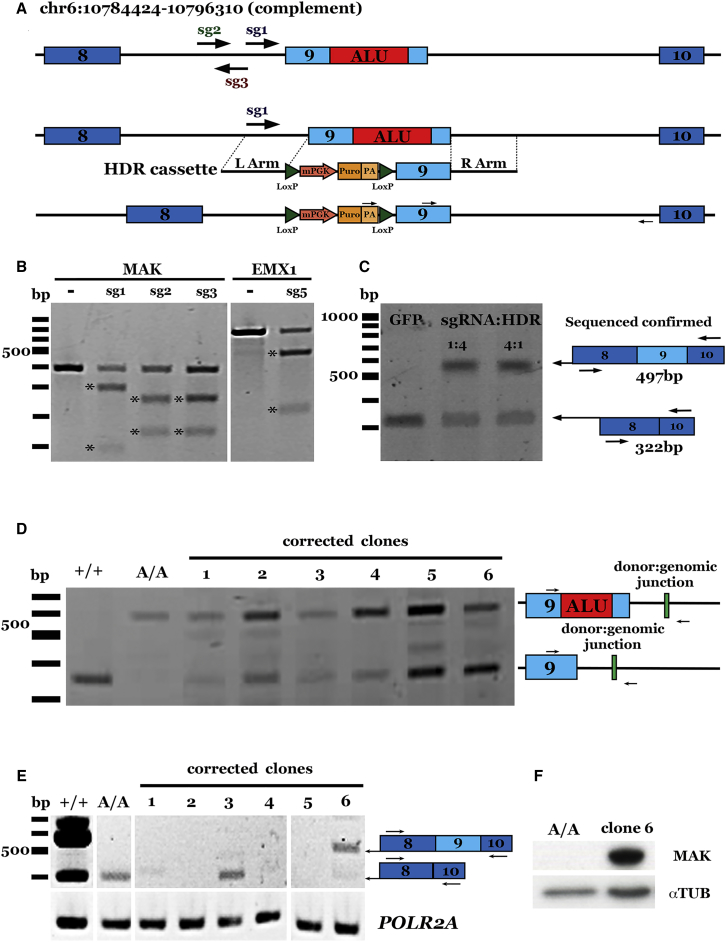
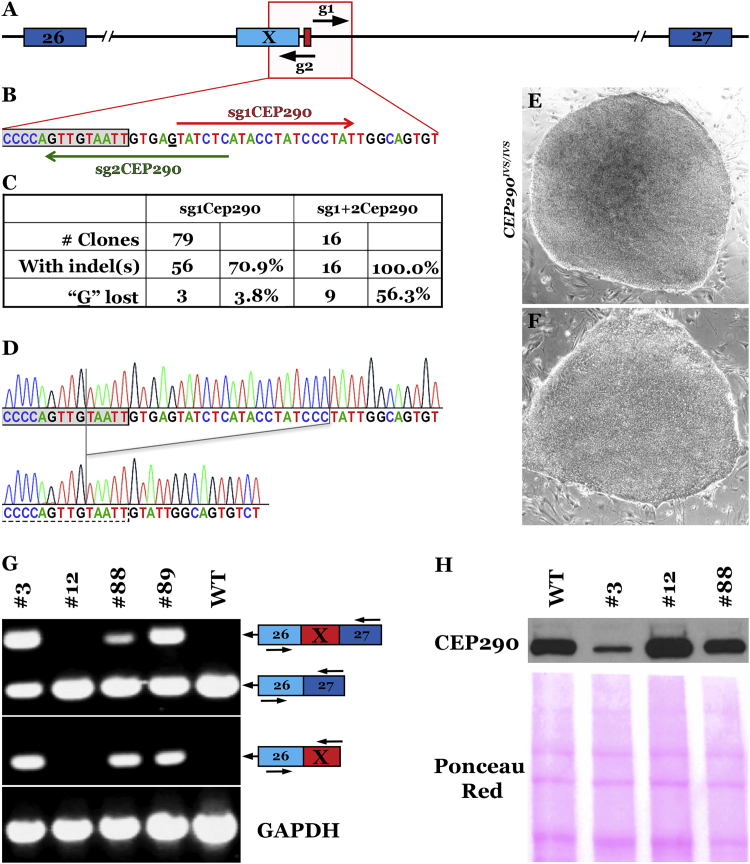
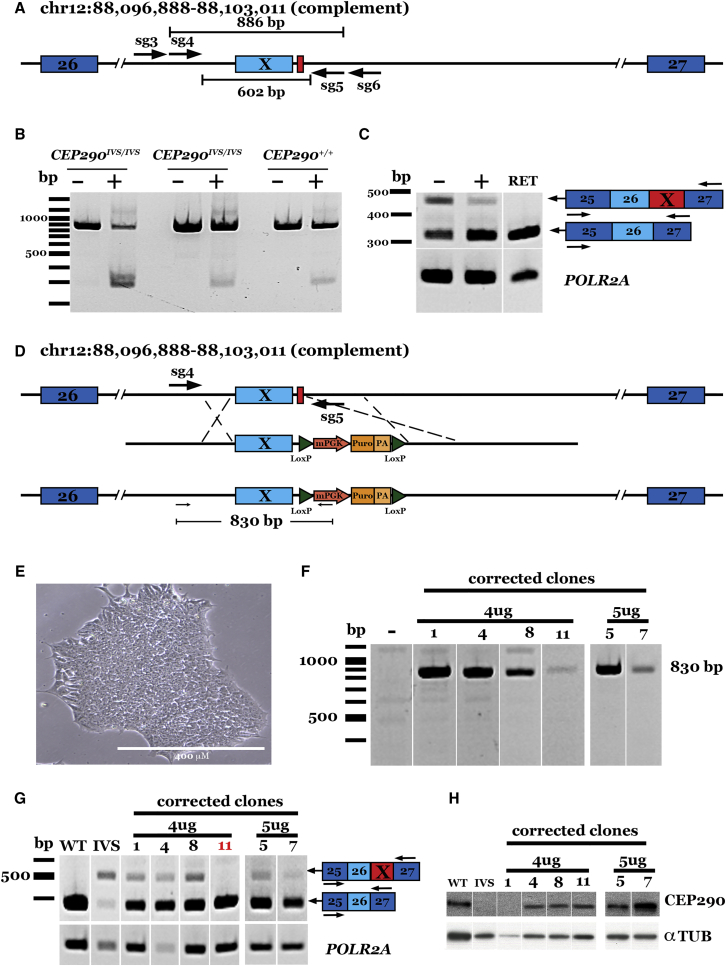
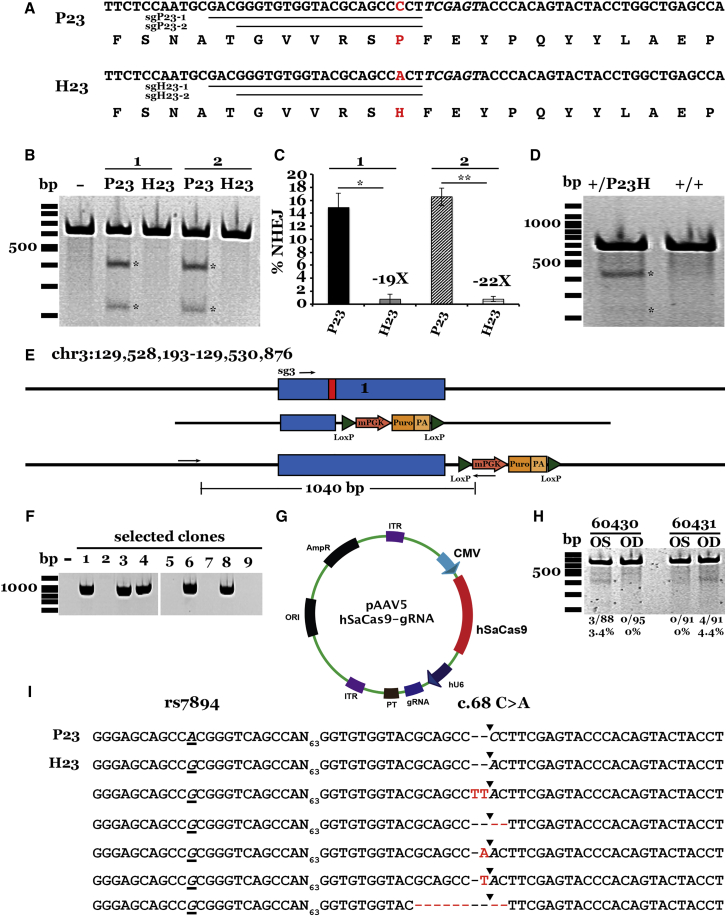
Patient-derived induced pluripotent stem cells (iPSCs) hold great promise for autologous cell replacement. However, for many inherited diseases, treatment will likely require genetic repair pre-transplantation. Genome editing technologies are useful for this application. The purpose of this study was to develop CRISPR-Cas9-mediated genome editing strategies to target and correct the three most common types of disease-causing variants in patient-derived iPSCs: (1) exonic, (2) deep intronic, and (3) dominant gain of function. We developed a homology-directed repair strategy targeting a homozygous Alu insertion in exon 9 of male germ cell-associated kinase (MAK) and demonstrated restoration of the retinal transcript and protein in patient cells. We generated a CRISPR-Cas9-mediated non-homologous end joining (NHEJ) approach to excise a major contributor to Leber congenital amaurosis, the IVS26 cryptic-splice mutation in CEP290, and demonstrated correction of the transcript and protein in patient iPSCs. Lastly, we designed allele-specific CRISPR guides that selectively target the mutant Pro23His rhodopsin (RHO) allele, which, following delivery to both patient iPSCs in vitro and pig retina in vivo, created a frameshift and premature stop that would prevent transcription of the disease-causing variant. The strategies developed in this study will prove useful for correcting a wide range of genetic variants in genes that cause inherited retinal degeneration.
Keywords: CRISPR; iPSCs; retinal degeneration.
Copyright © 2017 The American Society of Gene and Cell Therapy. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Gregory-Evans K., Bhattacharya S.S. Genetic blindness: current concepts in the pathogenesis of human outer retinal dystrophies. Trends Genet. 1998;14:103–108. - PubMed
-
- Bush R.A., Hawks K.W., Sieving P.A. Preservation of inner retinal responses in the aged Royal College of Surgeons rat. Evidence against glutamate excitotoxicity in photoreceptor degeneration. Invest. Ophthalmol. Vis. Sci. 1995;36:2054–2062. - PubMed
-
- Santos A., Humayun M.S., de Juan E., Jr., Greenburg R.J., Marsh M.J., Klock I.B., Milam A.H. Preservation of the inner retina in retinitis pigmentosa. A morphometric analysis. Arch. Ophthalmol. 1997;115:511–515. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
