

COP9 signalosome subunits protect Capicua from MAPK-dependent and -independent mechanisms of degradation
- PMID: 28619822
- PMCID: PMC5536928
- DOI: 10.1242/dev.148767
COP9 signalosome subunits protect Capicua from MAPK-dependent and -independent mechanisms of degradation
Abstract
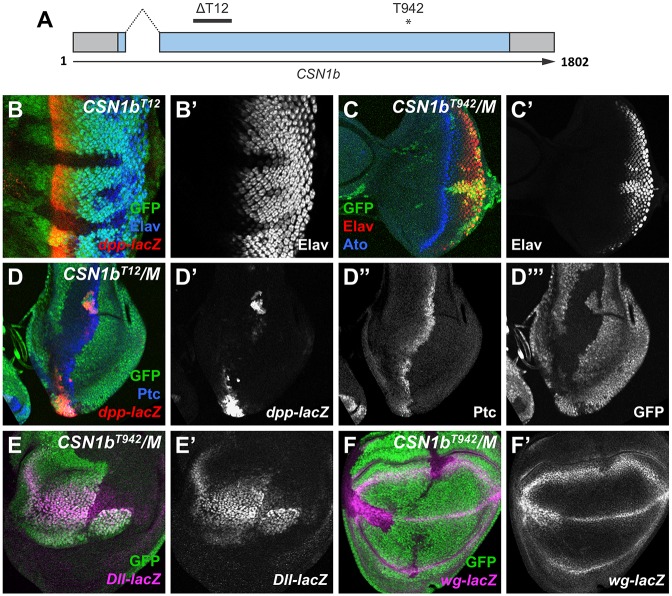
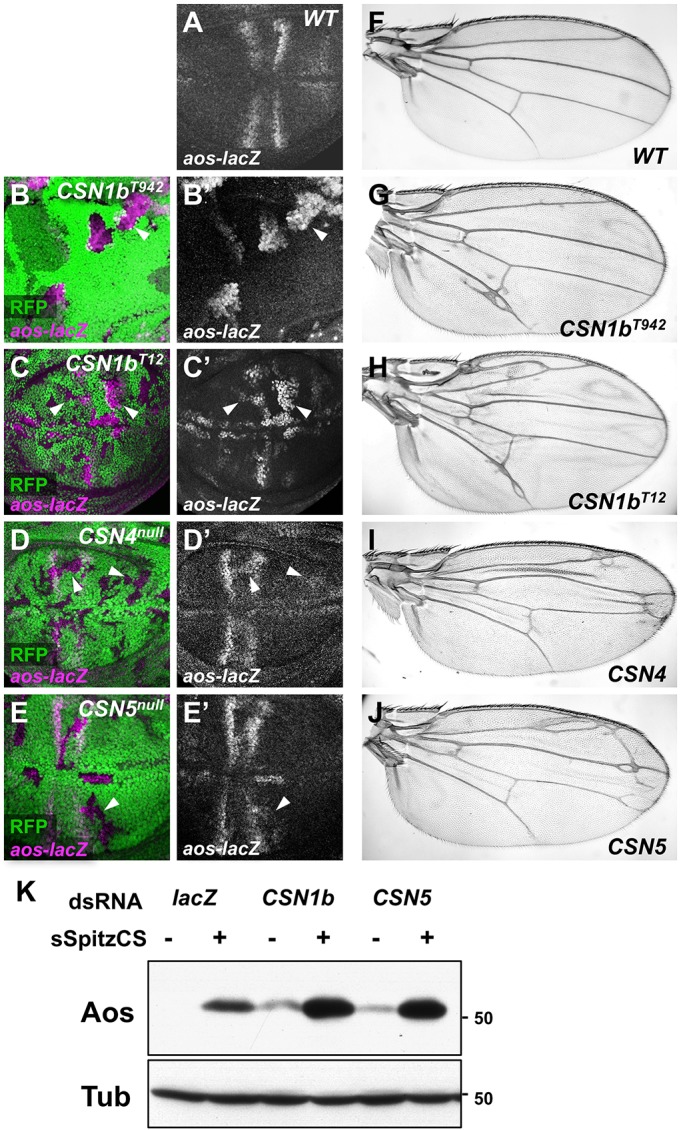
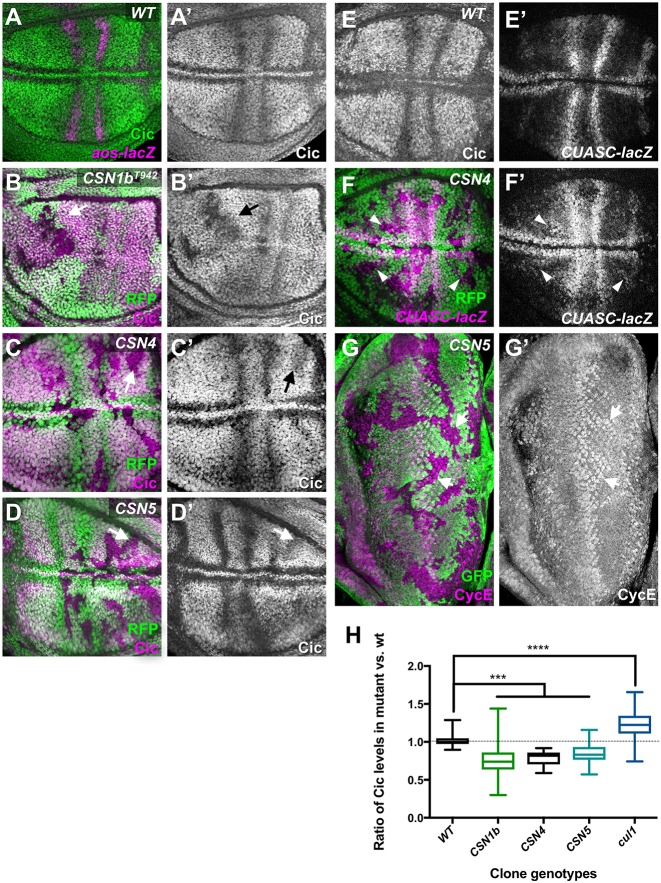
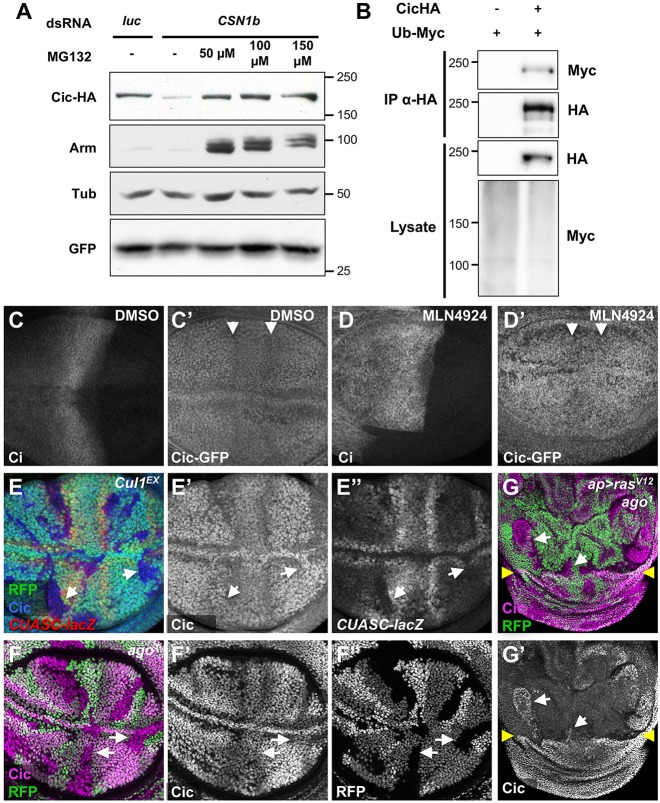
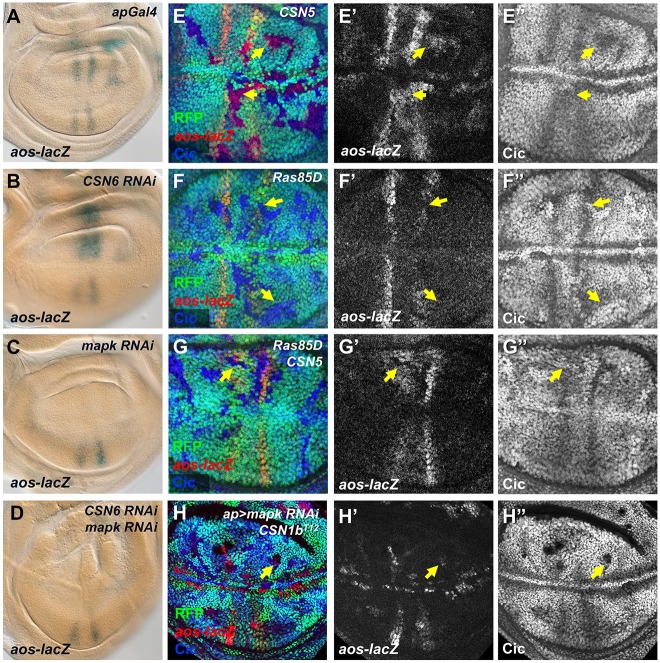
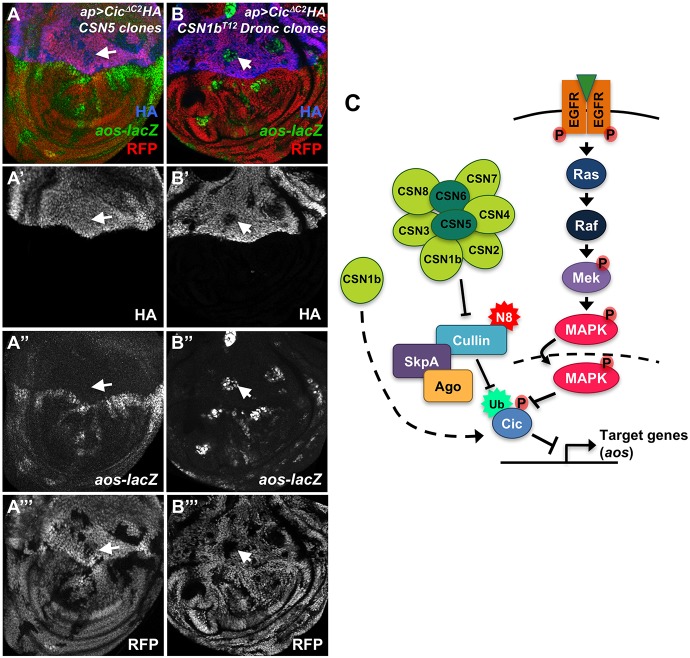
The COP9 signalosome removes Nedd8 modifications from the Cullin subunits of ubiquitin ligase complexes, reducing their activity. Here, we show that mutations in the Drosophila COP9 signalosome subunit 1b (CSN1b) gene increase the activity of ubiquitin ligases that contain Cullin 1. Analysis of CSN1b mutant phenotypes revealed a requirement for the COP9 signalosome to prevent ectopic expression of Epidermal growth factor receptor (EGFR) target genes. It does so by protecting Capicua, a transcriptional repressor of EGFR target genes, from EGFR pathway-dependent ubiquitylation by a Cullin 1/SKP1-related A/Archipelago E3 ligase and subsequent proteasomal degradation. The CSN1b subunit also maintains basal Capicua levels by protecting it from a separate mechanism of degradation that is independent of EGFR signaling. As a suppressor of tumor growth and metastasis, Capicua may be an important target of the COP9 signalosome in cancer.
Keywords: COP9 signalosome; Capicua; Drosophila melanogaster; EGFR; Nedd8; Ubiquitin ligase; Wing disc.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Beckmann E. A., Kohler A. M., Meister C., Christmann M., Draht O. W., Rakebrandt N., Valerius O. and Braus G. H. (2015). Integration of the catalytic subunit activates deneddylase activity in vivo as final step in fungal COP9 signalosome assembly. Mol. Microbiol. 97, 110-124. 10.1111/mmi.13017 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
