

Microbial Genetic Composition Tunes Host Longevity
- PMID: 28622510
- PMCID: PMC5635830
- DOI: 10.1016/j.cell.2017.05.036
Microbial Genetic Composition Tunes Host Longevity
Erratum in
-
Microbial Genetic Composition Tunes Host Longevity.Cell. 2018 May 3;173(4):1058. doi: 10.1016/j.cell.2018.04.026. Cell. 2018. PMID: 29727664 Free PMC article. No abstract available.
Abstract
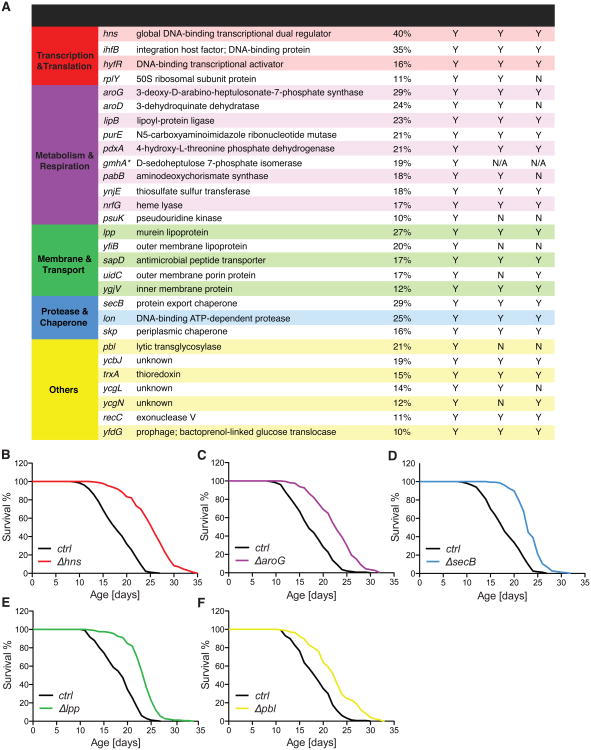
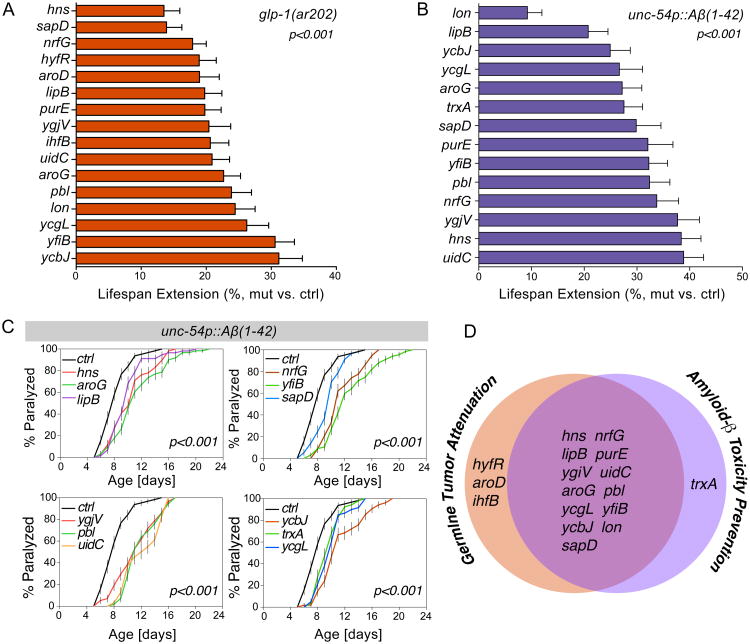
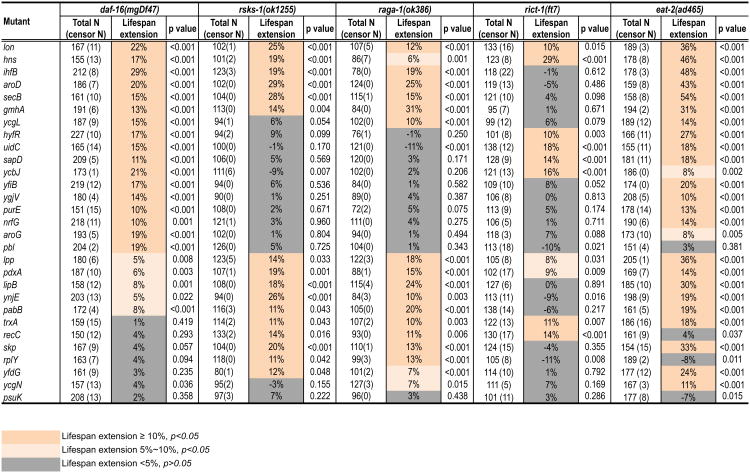
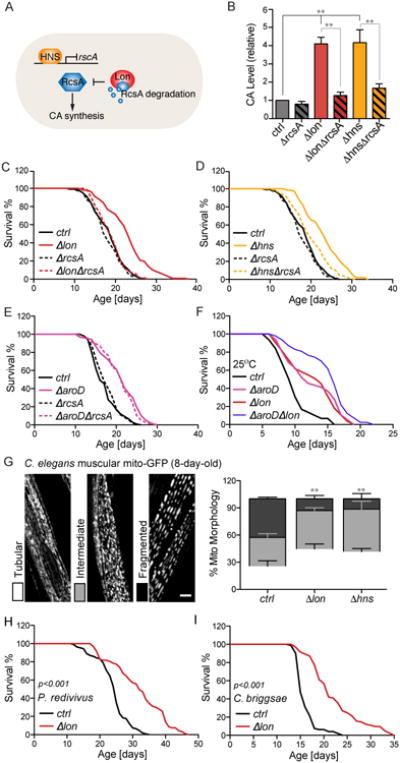
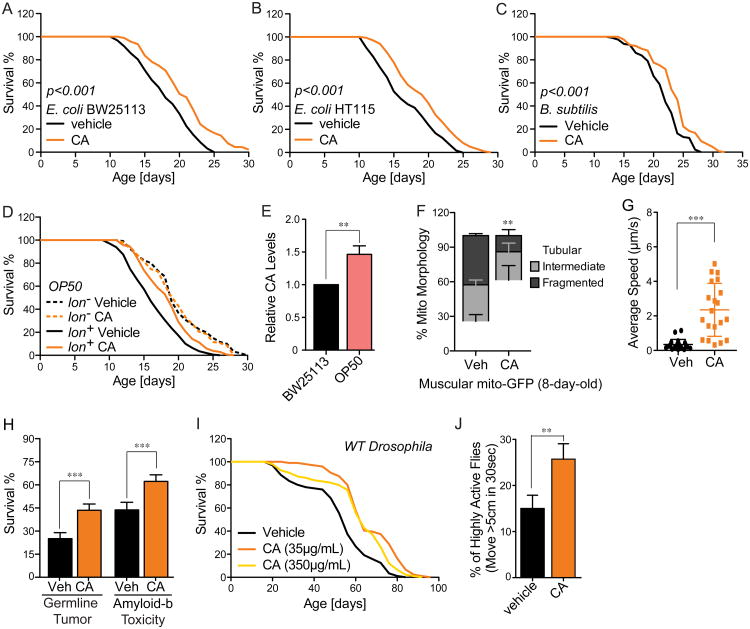
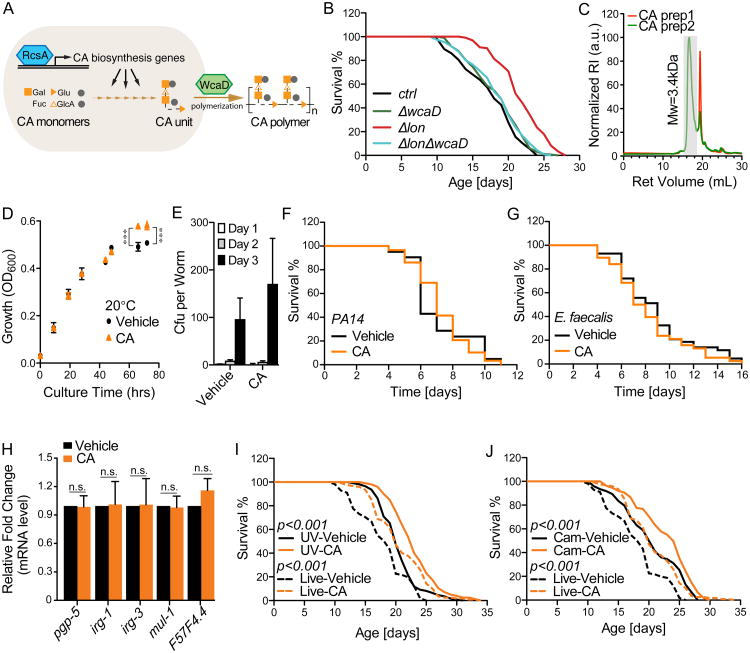
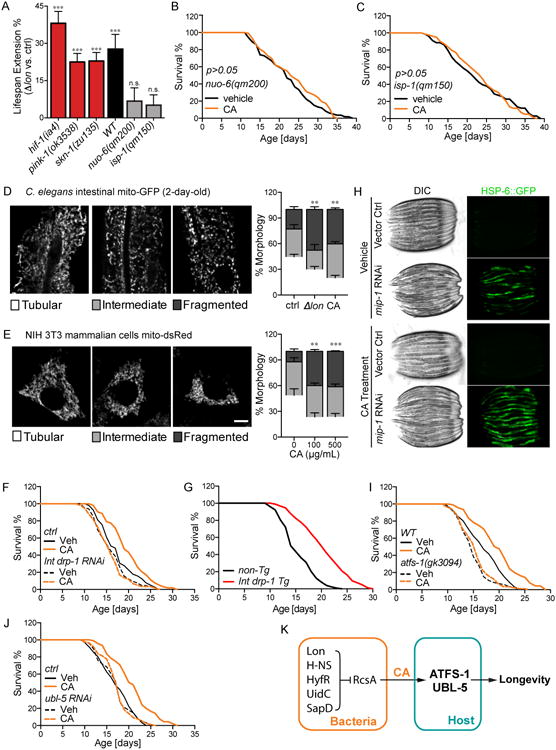
Homeostasis of the gut microbiota critically influences host health and aging. Developing genetically engineered probiotics holds great promise as a new therapeutic paradigm to promote healthy aging. Here, through screening 3,983 Escherichia coli mutants, we discovered that 29 bacterial genes, when deleted, increase longevity in the host Caenorhabditis elegans. A dozen of these bacterial mutants also protect the host from age-related progression of tumor growth and amyloid-beta accumulation. Mechanistically, we discovered that five bacterial mutants promote longevity through increased secretion of the polysaccharide colanic acid (CA), which regulates mitochondrial dynamics and unfolded protein response (UPRmt) in the host. Purified CA polymers are sufficient to promote longevity via ATFS-1, the host UPRmt-responsive transcription factor. Furthermore, the mitochondrial changes and longevity effects induced by CA are conserved across different species. Together, our results identified molecular targets for developing pro-longevity microbes and a bacterial metabolite acting on host mitochondria to promote longevity.
Keywords: colanic acid; longevity; microbiota-host interaction; mitochondrial dynamics; mitochondrial unfolded protein response; probiotics.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Microbiome and Longevity: Gut Microbes Send Signals to Host Mitochondria.Cell. 2017 Jun 15;169(7):1168-1169. doi: 10.1016/j.cell.2017.05.048. Cell. 2017. PMID: 28622501
References
-
- Andersson SG, Zomorodipour A, Andersson JO, Sicheritz-Ponten T, Alsmark UC, Podowski RM, Naslund AK, Eriksson AS, Winkler HH, Kurland CG. The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature. 1998;396:133–140. - PubMed
-
- Bentley R. The shikimate pathway--a metabolic tree with many branches. Critical reviews in biochemistry and molecular biology. 1990;25:307–384. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
