

Mir30c Is Involved in Diabetic Cardiomyopathy through Regulation of Cardiac Autophagy via BECN1
- PMID: 28624189
- PMCID: PMC5415963
- DOI: 10.1016/j.omtn.2017.03.005
Mir30c Is Involved in Diabetic Cardiomyopathy through Regulation of Cardiac Autophagy via BECN1
Abstract
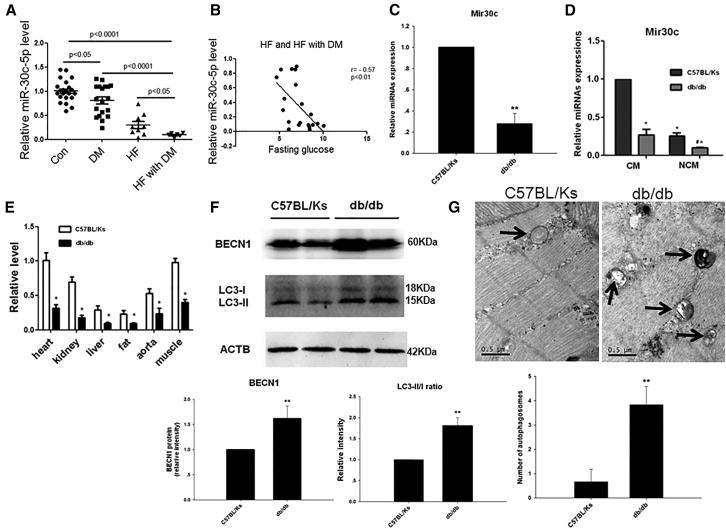
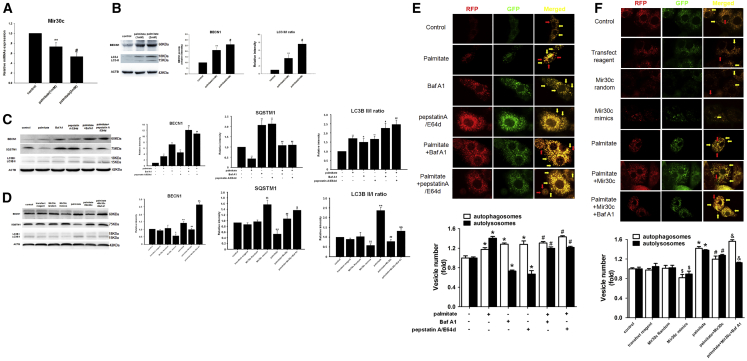
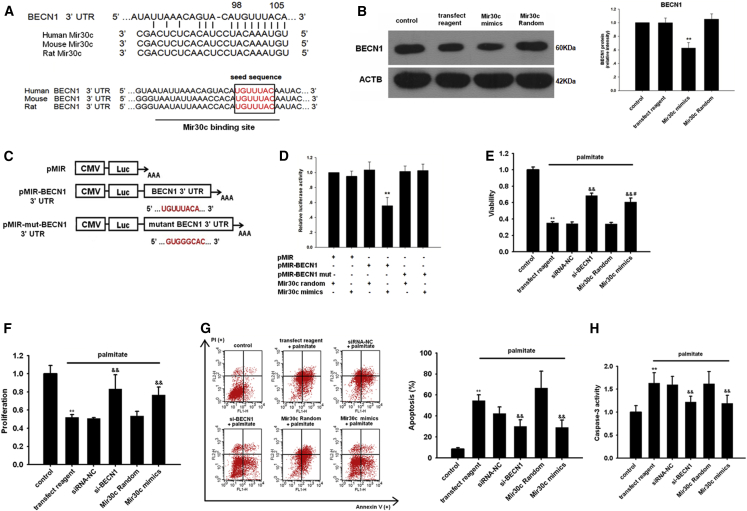
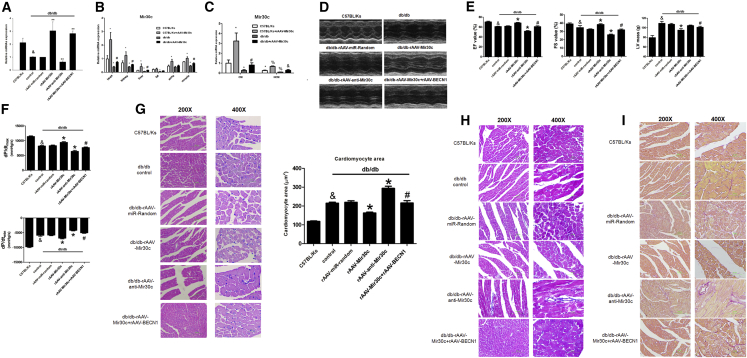
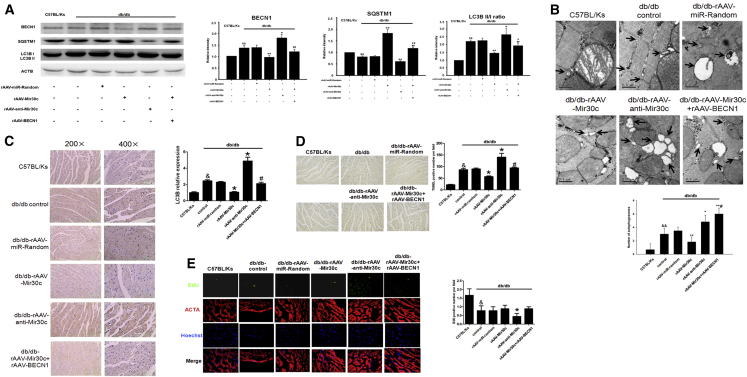
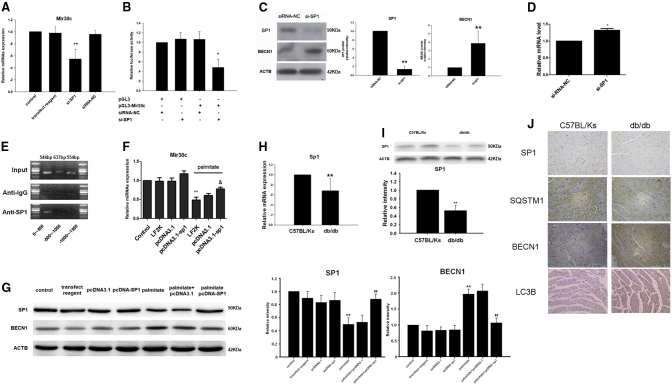
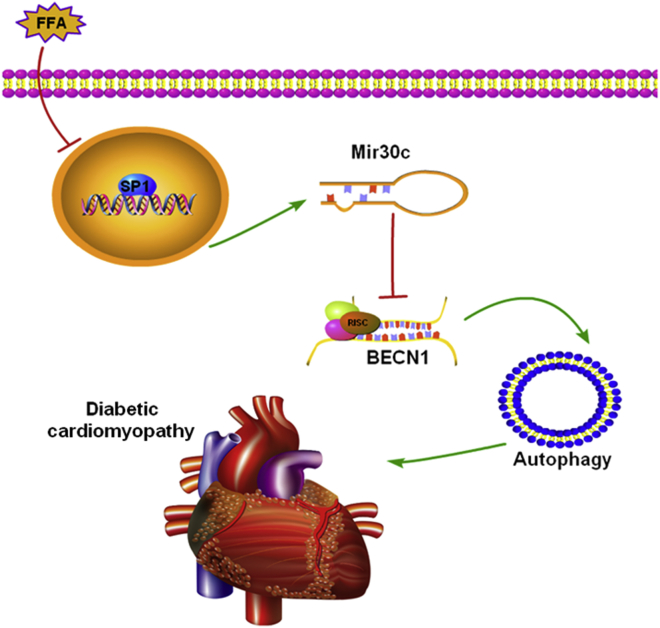
Multiple factors have been shown to promote the progression of diabetic cardiomyopathy. A link has previously been found between Mir30 and autophagy in cancer cells and in the heart, but the role of Mir30 in diabetic heart has not been studied. Using in vitro and in vivo approaches, we found that the depletion of Mir30c and induction of BECN1 enhanced autophagy in diabetic (db/db) hearts and in cardiomyocytes treated with the fatty acid palmitate. We verified that Mir30c repressed BECN1 expression by direct binding to the BECN1 3' UTRs. Mir30c overexpression inhibited the induction of BECN1 and subsequent autophagy in diabetic hearts and improved cardiac function and structure in diabetic mice. However, these effects were abrogated by BECN1 overexpression. Similarly, Mir30c knockdown resulted in increased BECN1 levels and autophagic flux, aggravating cardiac abnormalities. We also show that SP1, an important transcriptional factor in energy metabolism regulation, is a key upstream activator of Mir30c that binds the promoter region of Mir30c. Our findings indicate that downregulation of Mir30c and subsequent activation of BECN1 promotes autophagy, contributing to the pathogenesis of diabetic cardiomyopathy. This observation suggests a theoretical ground for developing microRNA-based therapeutics against diabetic cardiomyopathy by inhibiting autophagy.
Keywords: BECN1; Mir30c; autophagy; cardiac dysfunction; diabetes.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Rubler S., Dlugash J., Yuceoglu Y.Z., Kumral T., Branwood A.W., Grishman A. New type of cardiomyopathy associated with diabetic glomerulosclerosis. Am. J. Cardiol. 1972;30:595–602. - PubMed
-
- Boudina S., Abel E.D. Diabetic cardiomyopathy revisited. Circulation. 2007;115:3213–3223. - PubMed
-
- Belke D.D., Larsen T.S., Gibbs E.M., Severson D.L. Altered metabolism causes cardiac dysfunction in perfused hearts from diabetic (db/db) mice. Am. J. Physiol. Endocrinol. Metab. 2000;279:E1104–E1113. - PubMed
-
- Boudina S., Sena S., O’Neill B.T., Tathireddy P., Young M.E., Abel E.D. Reduced mitochondrial oxidative capacity and increased mitochondrial uncoupling impair myocardial energetics in obesity. Circulation. 2005;112:2686–2695. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
