

A Corticothalamic Circuit for Dynamic Switching between Feature Detection and Discrimination
- PMID: 28625486
- PMCID: PMC5568886
- DOI: 10.1016/j.neuron.2017.05.019
A Corticothalamic Circuit for Dynamic Switching between Feature Detection and Discrimination
Abstract
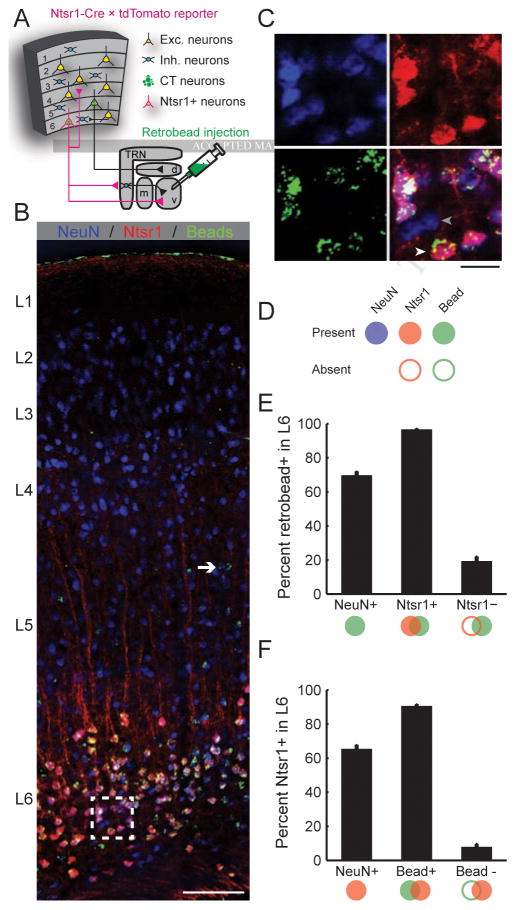
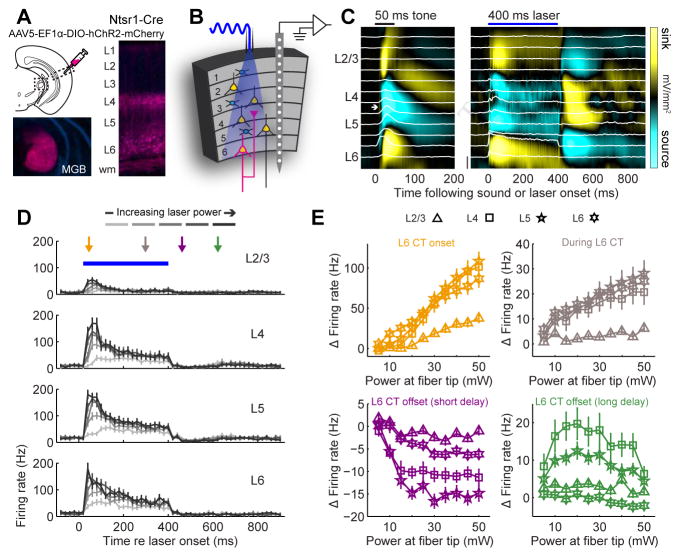
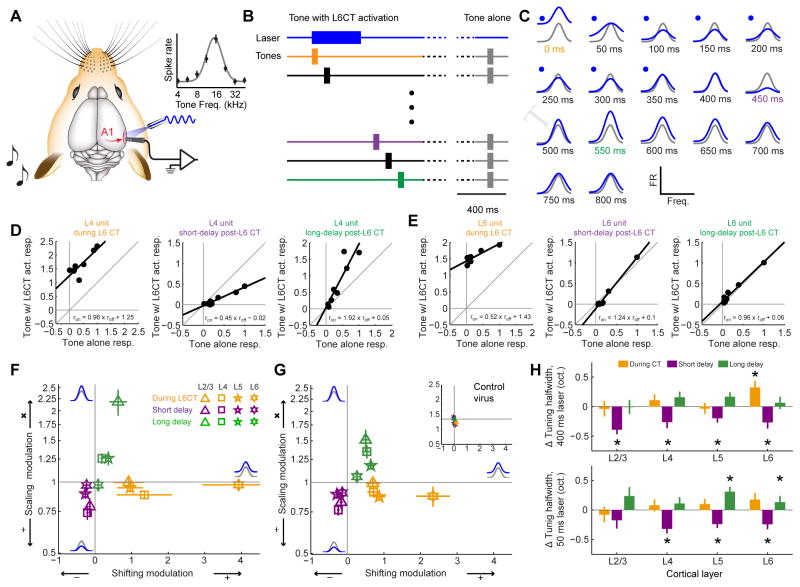
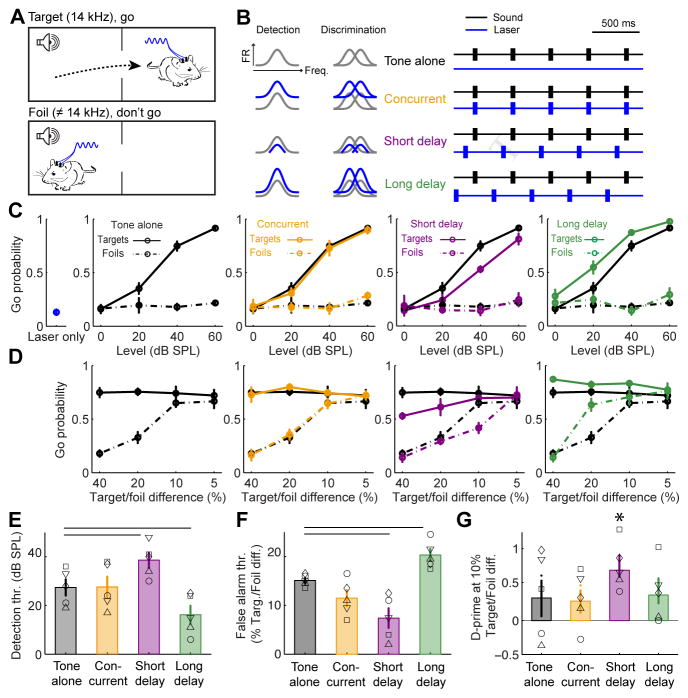
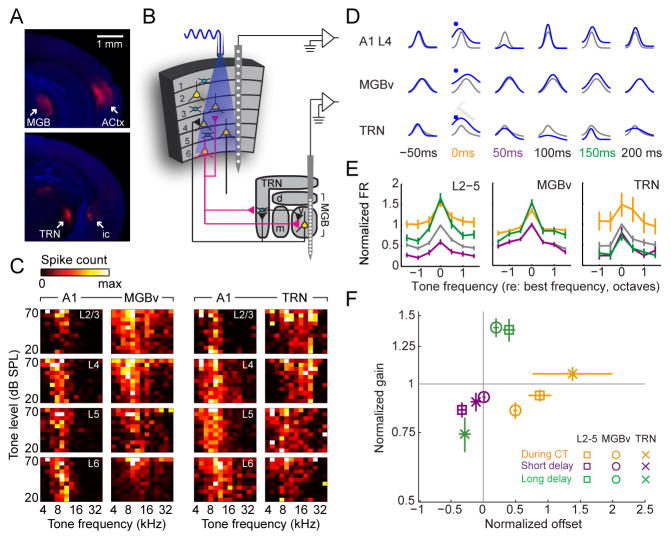
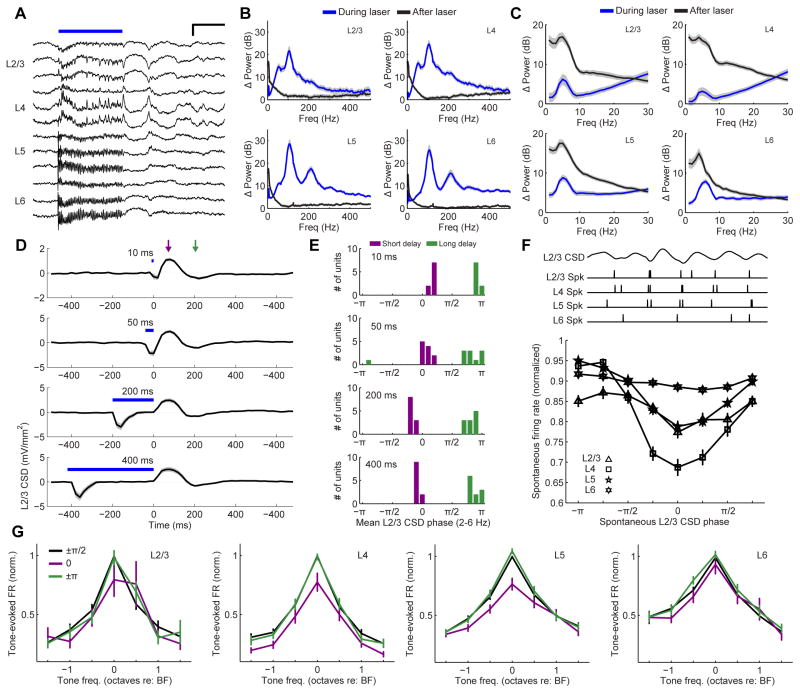
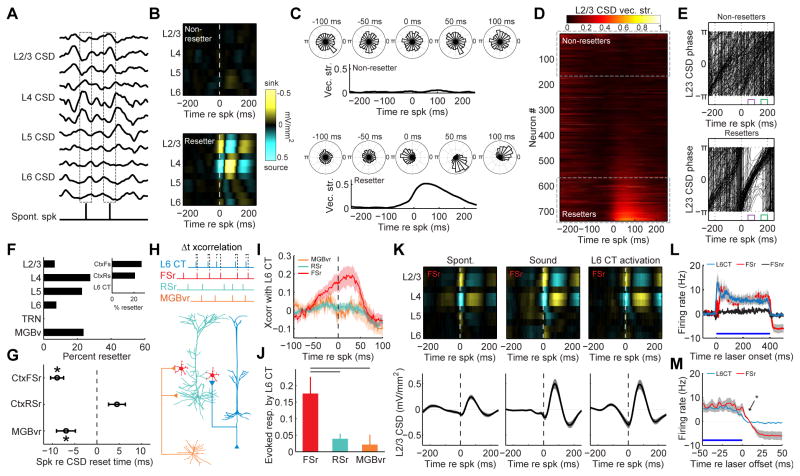
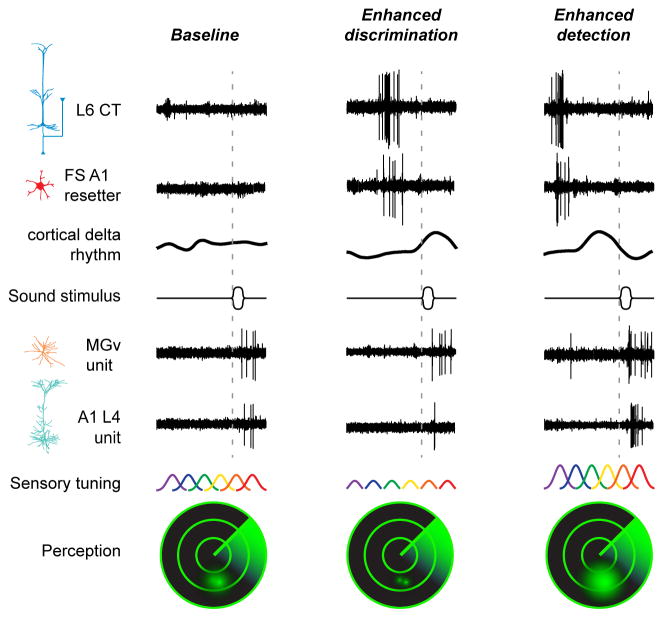
Sensory processing must be sensitive enough to encode faint signals near the noise floor but selective enough to differentiate between similar stimuli. Here we describe a layer 6 corticothalamic (L6 CT) circuit in the mouse auditory forebrain that alternately biases sound processing toward hypersensitivity and improved behavioral sound detection or dampened excitability and enhanced sound discrimination. Optogenetic activation of L6 CT neurons could increase or decrease the gain and tuning precision in the thalamus and all layers of the cortical column, depending on the timing between L6 CT activation and sensory stimulation. The direction of neural and perceptual modulation - enhanced detection at the expense of discrimination or vice versa - arose from the interaction of L6 CT neurons and subnetworks of fast-spiking inhibitory neurons that reset the phase of low-frequency cortical rhythms. These findings suggest that L6 CT neurons contribute to the resolution of the competing demands of detection and discrimination.
Keywords: auditory cortex; auditory thalamus; delta rhythm; layer 6; medial geniculate body; modulation; oscillation; phase reset; plasticity; theta rhythm.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Bourassa J, Deschenes M. Corticothalamic projections from the primary visual cortex in rats: a single-fiber study using biocytin as an anterograde tracer. Eur J Neurosci. 1995;7:19–30. - PubMed
STAR Methods References
-
- Kaur S, Rose HJ, Lazar R, Liang K, Metherate R. Spectral integration in primary auditory cortex: Laminar processing of afferent input, in vivo and in vitro. Neuroscience. 2005;134:1033–1045. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
