

BACE1 Cleavage Site Selection Critical for Amyloidogenesis and Alzheimer's Pathogenesis
- PMID: 28626014
- PMCID: PMC6705717
- DOI: 10.1523/JNEUROSCI.0340-17.2017
BACE1 Cleavage Site Selection Critical for Amyloidogenesis and Alzheimer's Pathogenesis
Abstract
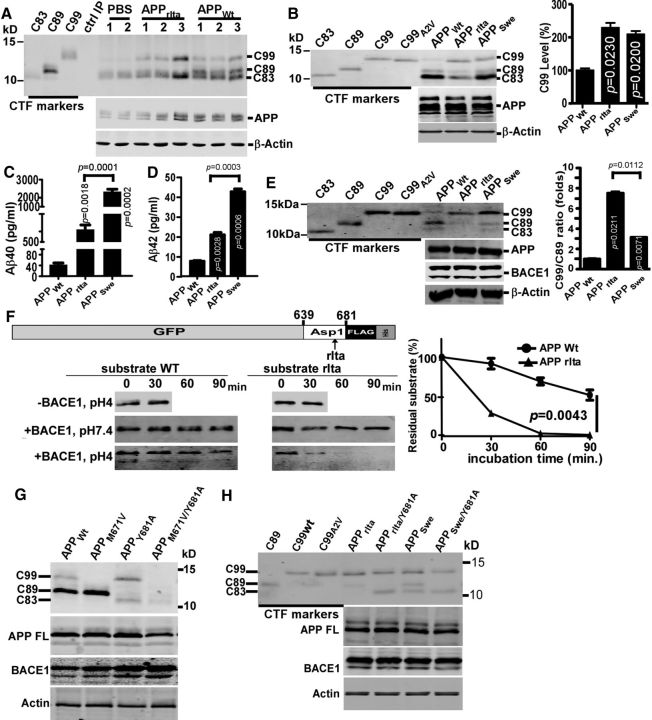
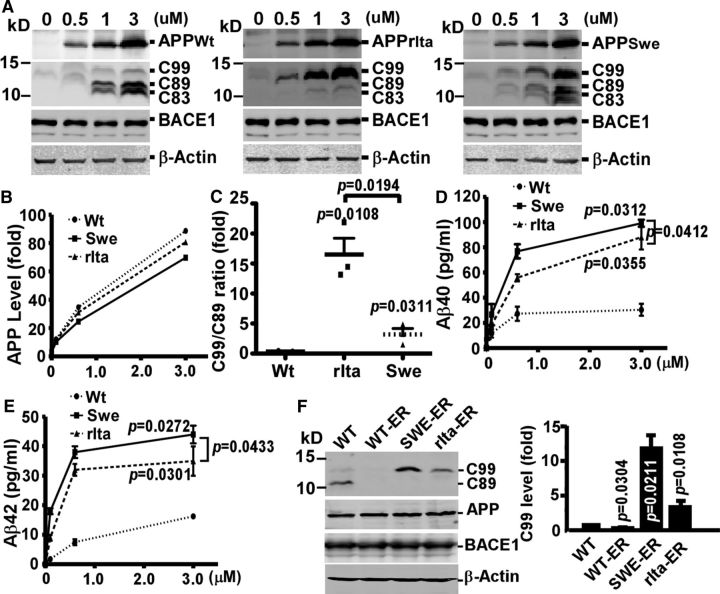
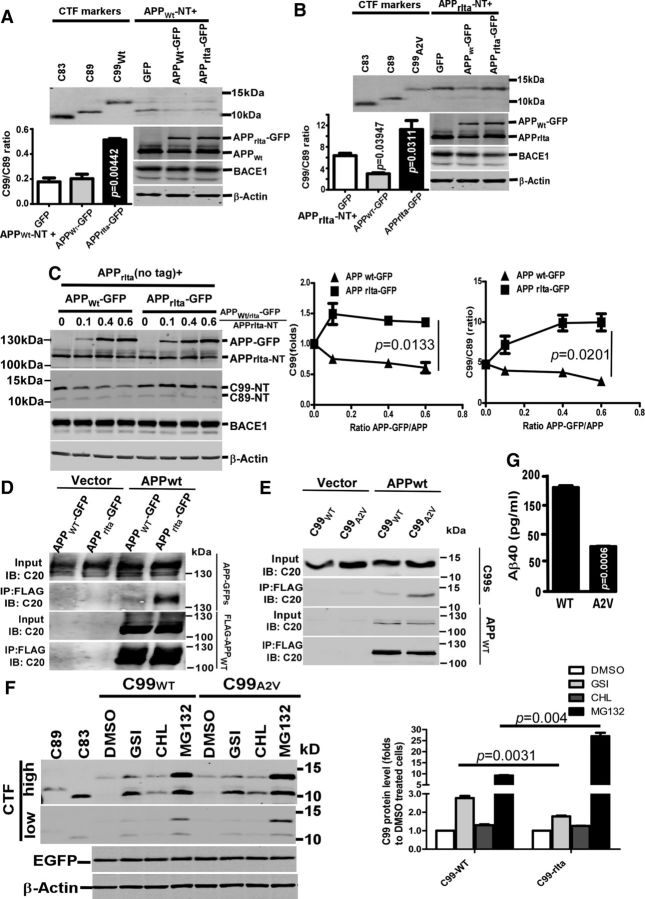
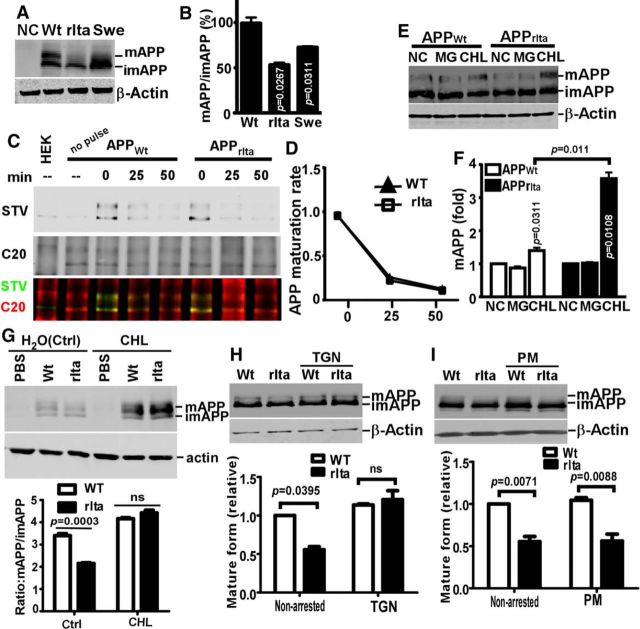
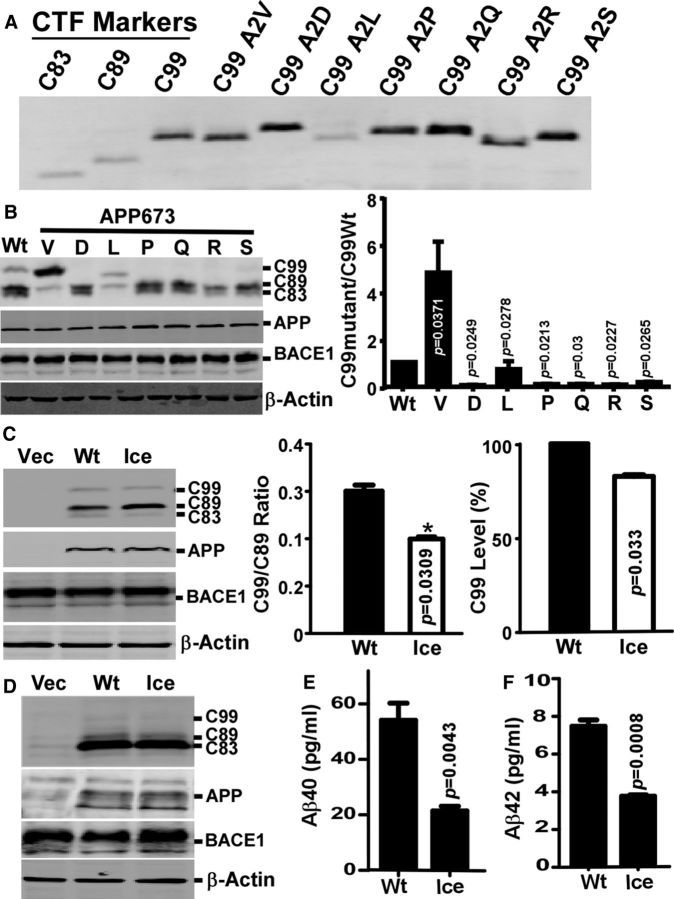
Mutations in amyloid β precursor protein (APP) gene alter APP processing, either causing familial Alzheimer's disease (AD) or protecting against dementia. Under normal conditions, β-site APP cleaving enzyme 1 (BACE1) cleaves APP at minor Asp1 site to generate C99 for amyloid β protein (Aβ) production, and predominantly at major Glu11 site to generate C89, resulting in truncated Aβ production. We discovered that A673V mutation, the only recessive AD-associated APP mutation, shifted the preferential β-cleavage site of BACE1 in APP from the Glu11 site to the Asp1 site both in male and female transgenic mice in vivo and in cell lines and primary neuronal culture derived from timed pregnant rats in vitro, resulting in a much higher C99 level and C99/C89 ratio. All other mutations at this site, including the protective Icelandic A673T mutation, reduced C99 generation, and decreased the C99/C89 ratio. Furthermore, A673V mutation caused stronger dimerization between mutant and wild-type APP, enhanced the lysosomal degradation of the mutant APP, and inhibited γ-secretase cleavage of the mutant C99 to generate Aβ, leading to recessively inherited AD. The results demonstrate that APP673 regulates APP processing and the BACE1 cleavage site selection is critical for amyloidogenesis in AD pathogenesis, and implicate a pharmaceutical potential for targeting the APP673 site for AD drug development.SIGNIFICANCE STATEMENT β-site APP cleaving enzyme 1 (BACE1) is essential for amyloid β protein production. We discovered that A673V mutation shifted the BACE1 cleavage site from the Glu11 to the Asp1 site, resulting in much higher C99 level and C99/C89 ratio. All other mutations at this site of amyloid β precursor protein (APP) reduced C99 generation and decreased the C99/C89 ratio. Furthermore, A673V mutation resulted in stronger dimerization between mutant and wild-type APP, enhanced the lysosomal degradation of the mutant APP, and inhibited γ-secretase cleavage of the mutant C99 to generate amyloid β protein, leading to recessively inherited Alzheimer's disease (AD). The results demonstrate that APP673 regulates APP processing, and the BACE1 cleavage site selection is critical for amyloidogenesis in AD pathogenesis, and implicate a pharmaceutical potential for targeting the APP673 site for AD drug development.
Keywords: Alzheimer's disease; BACE1.
Copyright © 2017 the authors 0270-6474/17/376915-11$15.00/0.
Figures
Comment in
-
New Insights on the Role of Residue 673 of APP in Alzheimer's Disease.J Neurosci. 2018 Jan 17;38(3):515-517. doi: 10.1523/JNEUROSCI.2710-17.2017. J Neurosci. 2018. PMID: 29343592 Free PMC article. No abstract available.
References
-
- Benilova I, Gallardo R, Ungureanu AA, Castillo Cano V, Snellinx A, Ramakers M, Bartic C, Rousseau F, Schymkowitz J, De Strooper B (2014) The Alzheimer disease protective mutation A2T modulates kinetic and thermodynamic properties of amyloid-beta (Abeta) aggregation. J Biol Chem 289:30977–30989. 10.1074/jbc.M114.599027 - DOI - PMC - PubMed
-
- De Jonghe C, Esselens C, Kumar-Singh S, Craessaerts K, Serneels S, Checler F, Annaert W, Van Broeckhoven C, De Strooper B (2001) Pathogenic APP mutations near the gamma-secretase cleavage site differentially affect Abeta secretion and APP C-terminal fragment stability. Hum Mol Genet 10:1665–1671. 10.1093/hmg/10.16.1665 - DOI - PubMed
-
- Deng Y, Wang Z, Wang R, Zhang X, Zhang S, Wu Y, Staufenbiel M, Cai F, Song W (2013) Amyloid-beta protein (Abeta) Glu11 is the major beta-secretase site of beta-site amyloid-beta precursor protein-cleaving enzyme 1(BACE1), and shifting the cleavage site to Abeta Asp1 contributes to Alzheimer pathogenesis. Eur J Neurosci 37:1962–1969. 10.1111/ejn.12235 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical