

The Diverse Roles of Arrestin Scaffolds in G Protein-Coupled Receptor Signaling
- PMID: 28626043
- PMCID: PMC5482185
- DOI: 10.1124/pr.116.013367
The Diverse Roles of Arrestin Scaffolds in G Protein-Coupled Receptor Signaling
Abstract
The visual/β-arrestins, a small family of proteins originally described for their role in the desensitization and intracellular trafficking of G protein-coupled receptors (GPCRs), have emerged as key regulators of multiple signaling pathways. Evolutionarily related to a larger group of regulatory scaffolds that share a common arrestin fold, the visual/β-arrestins acquired the capacity to detect and bind activated GPCRs on the plasma membrane, which enables them to control GPCR desensitization, internalization, and intracellular trafficking. By acting as scaffolds that bind key pathway intermediates, visual/β-arrestins both influence the tonic level of pathway activity in cells and, in some cases, serve as ligand-regulated scaffolds for GPCR-mediated signaling. Growing evidence supports the physiologic and pathophysiologic roles of arrestins and underscores their potential as therapeutic targets. Circumventing arrestin-dependent GPCR desensitization may alleviate the problem of tachyphylaxis to drugs that target GPCRs, and find application in the management of chronic pain, asthma, and psychiatric illness. As signaling scaffolds, arrestins are also central regulators of pathways controlling cell growth, migration, and survival, suggesting that manipulating their scaffolding functions may be beneficial in inflammatory diseases, fibrosis, and cancer. In this review we examine the structure-function relationships that enable arrestins to perform their diverse roles, addressing arrestin structure at the molecular level, the relationship between arrestin conformation and function, and sites of interaction between arrestins, GPCRs, and nonreceptor-binding partners. We conclude with a discussion of arrestins as therapeutic targets and the settings in which manipulating arrestin function might be of clinical benefit.
Copyright © 2017 by The American Society for Pharmacology and Experimental Therapeutics.
Figures
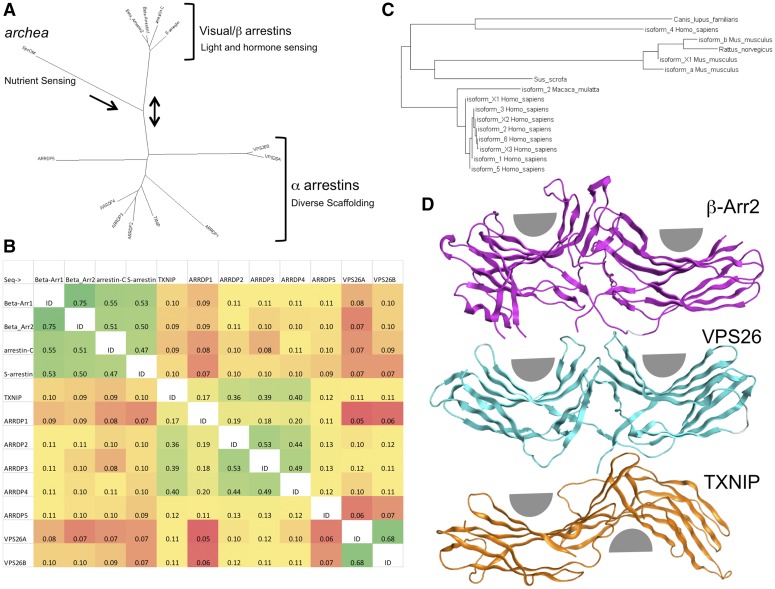

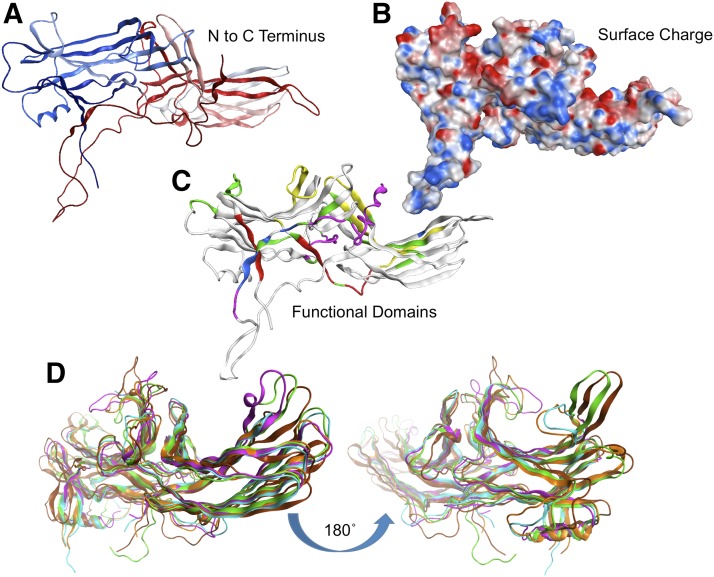
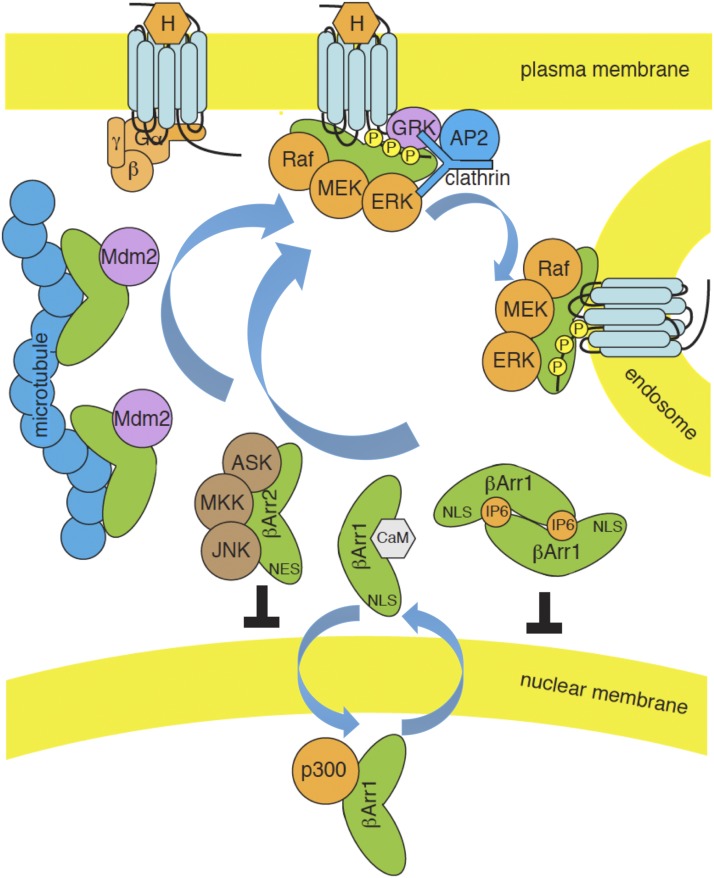
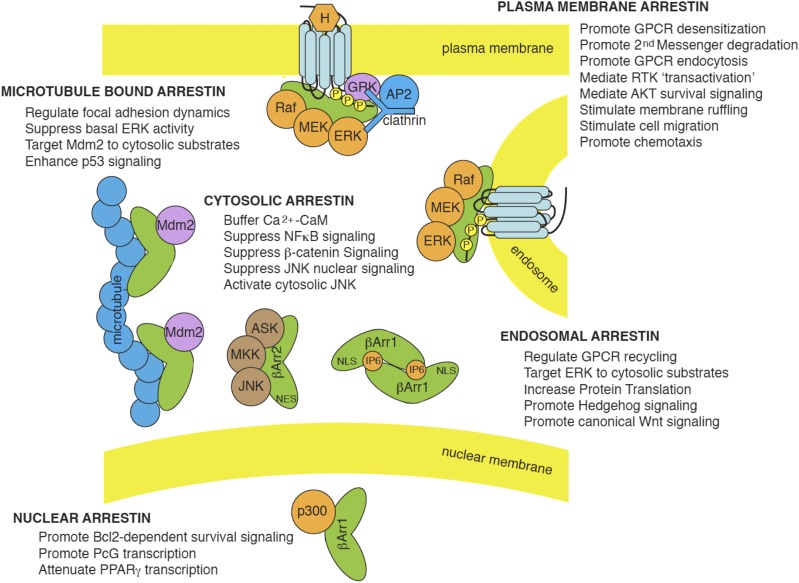
References
-
- Abraham D, Podar K, Pacher M, Kubicek M, Welzel N, Hemmings BA, Dilworth SM, Mischak H, Kolch W, Baccarini M. (2000) Raf-1-associated protein phosphatase 2A as a positive regulator of kinase activation. J Biol Chem 275:22300–22304. - PubMed
-
- Abrahamsen H, Baillie G, Ngai J, Vang T, Nika K, Ruppelt A, Mustelin T, Zaccolo M, Houslay M, Taskén K. (2004) TCR- and CD28-mediated recruitment of phosphodiesterase 4 to lipid rafts potentiates TCR signaling. J Immunol 173:4847–4858. - PubMed
-
- Adams DG, Coffee RL, Jr, Zhang H, Pelech S, Strack S, Wadzinski BE. (2005) Positive regulation of Raf1-MEK1/2-ERK1/2 signaling by protein serine/threonine phosphatase 2A holoenzymes. J Biol Chem 280:42644–42654. - PubMed
-
- Ahlquist RP. (1948) A study of the adrenotropic receptors. Am J Physiol 153:586–600. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
