

The Lung is a Host Defense Niche for Immediate Neutrophil-Mediated Vascular Protection
- PMID: 28626833
- PMCID: PMC5472445
- DOI: 10.1126/sciimmunol.aam8929
The Lung is a Host Defense Niche for Immediate Neutrophil-Mediated Vascular Protection
Abstract
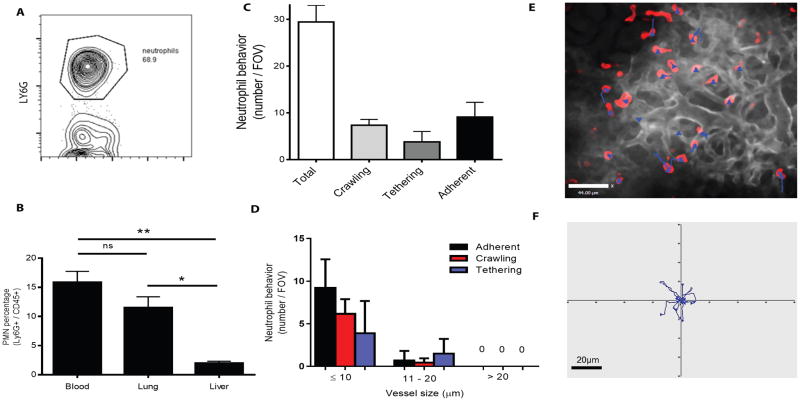
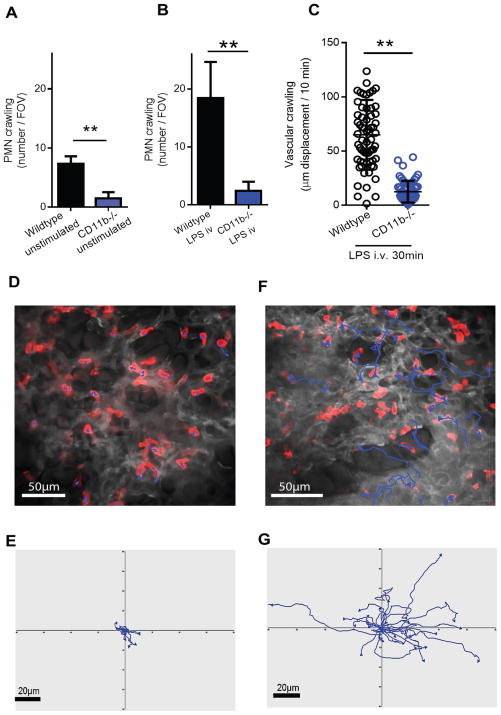
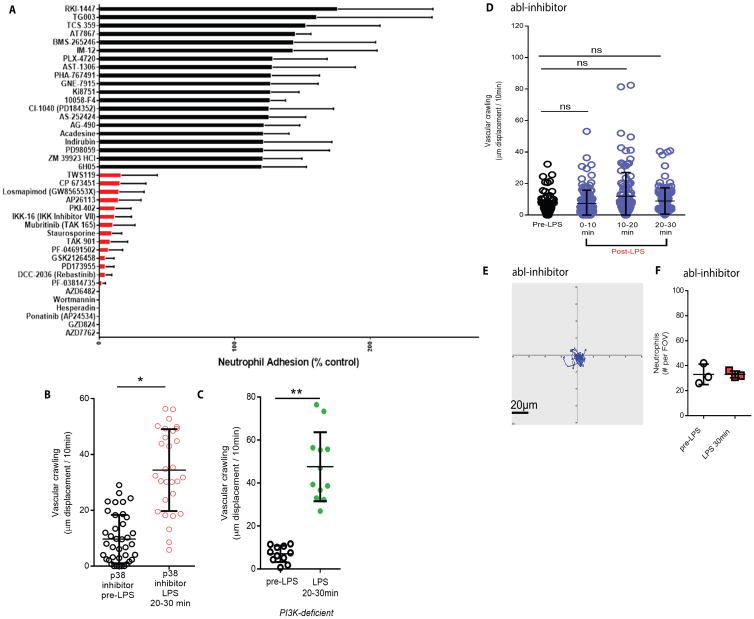
Bloodstream infection is a hallmark of sepsis, a medically emergent condition requiring rapid treatment. However, upregulation of host defense proteins through toll-like receptors and NFκB requires hours after endotoxin detection. Using confocal pulmonary intravital microscopy, we identified that the lung provides a TLR4-Myd88-and abl tyrosine kinase-dependent niche for immediate CD11b-dependent neutrophil responses to endotoxin and Gram-negative bloodstream pathogens. In an in vivo model of bacteremia, neutrophils crawled to and rapidly phagocytosed Escherichia coli sequestered to the lung endothelium. Therefore, the lung capillaries provide a vascular defensive niche whereby endothelium and neutrophils cooperate for immediate detection and capture of disseminating pathogens.
Conflict of interest statement
Competing interests: The authors declare no competing interests
Figures





References
-
- Liu V, Escobar GJ, Greene JD, Soule J, Whippy A, Angus DC, Iwashyna TJ. Hospital deaths in patients with sepsis from 2 independent cohorts. JAMA. 2014;312:90–92. - PubMed
-
- Laupland KB. Incidence of bloodstream infection: a review of population-based studies. Clin Microbiol Infect. 2013;19:492–500. - PubMed
-
- Biondi E, Evans R, Mischler M, Bendel-Stenzel M, Horstmann S, Lee V, Aldag J, Gigliotti F. Epidemiology of bacteremia in febrile infants in the United States. Pediatrics. 2013;132:990–996. - PubMed
-
- Deen J, von Seidlein L, Andersen F, Elle N, White NJ, Lubell Y. Community-acquired bacterial bloodstream infections in developing countries in south and southeast Asia: a systematic review. Lancet Infect Dis. 2012;12:480–487. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
