

HBV RNA pre-genome encodes specific motifs that mediate interactions with the viral core protein that promote nucleocapsid assembly
- PMID: 28628133
- PMCID: PMC5495169
- DOI: 10.1038/nmicrobiol.2017.98
HBV RNA pre-genome encodes specific motifs that mediate interactions with the viral core protein that promote nucleocapsid assembly
Abstract
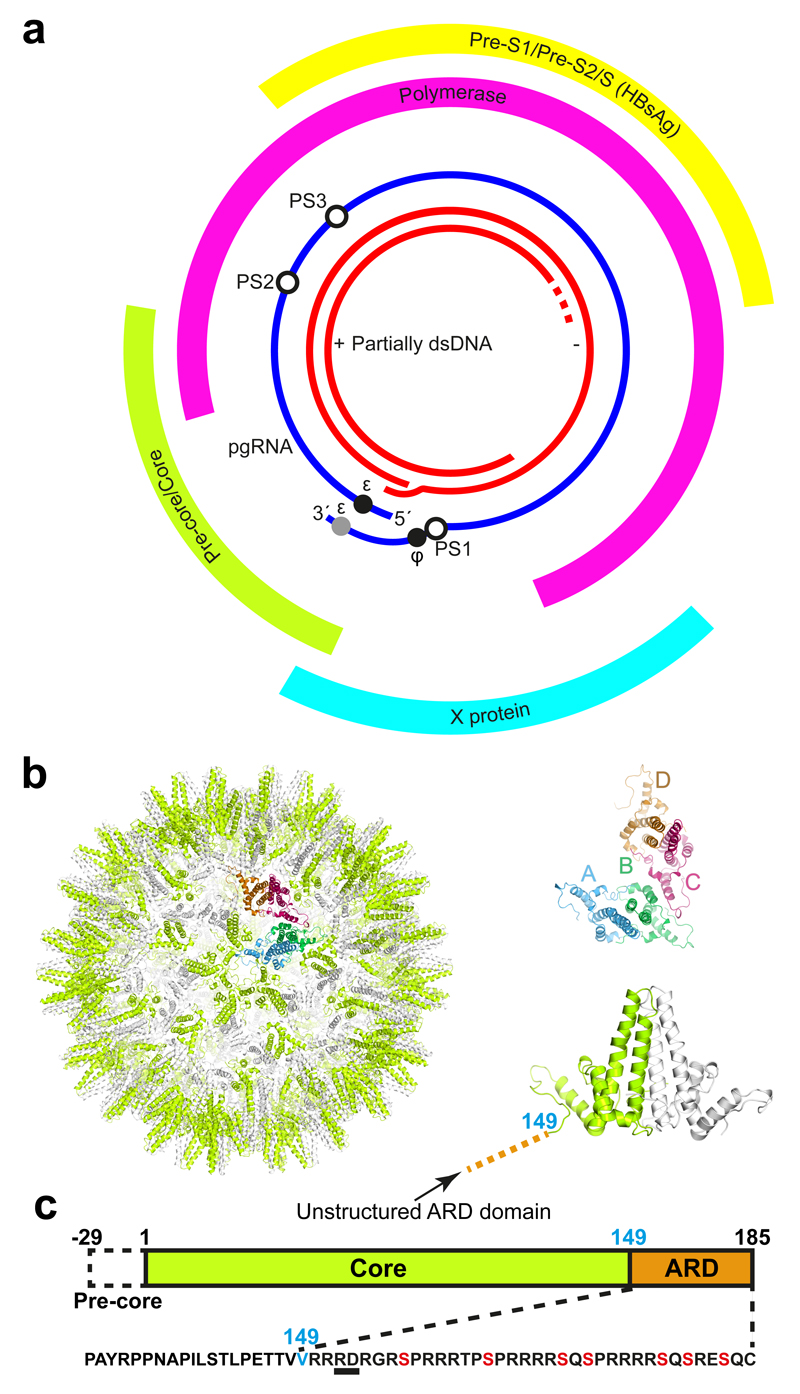
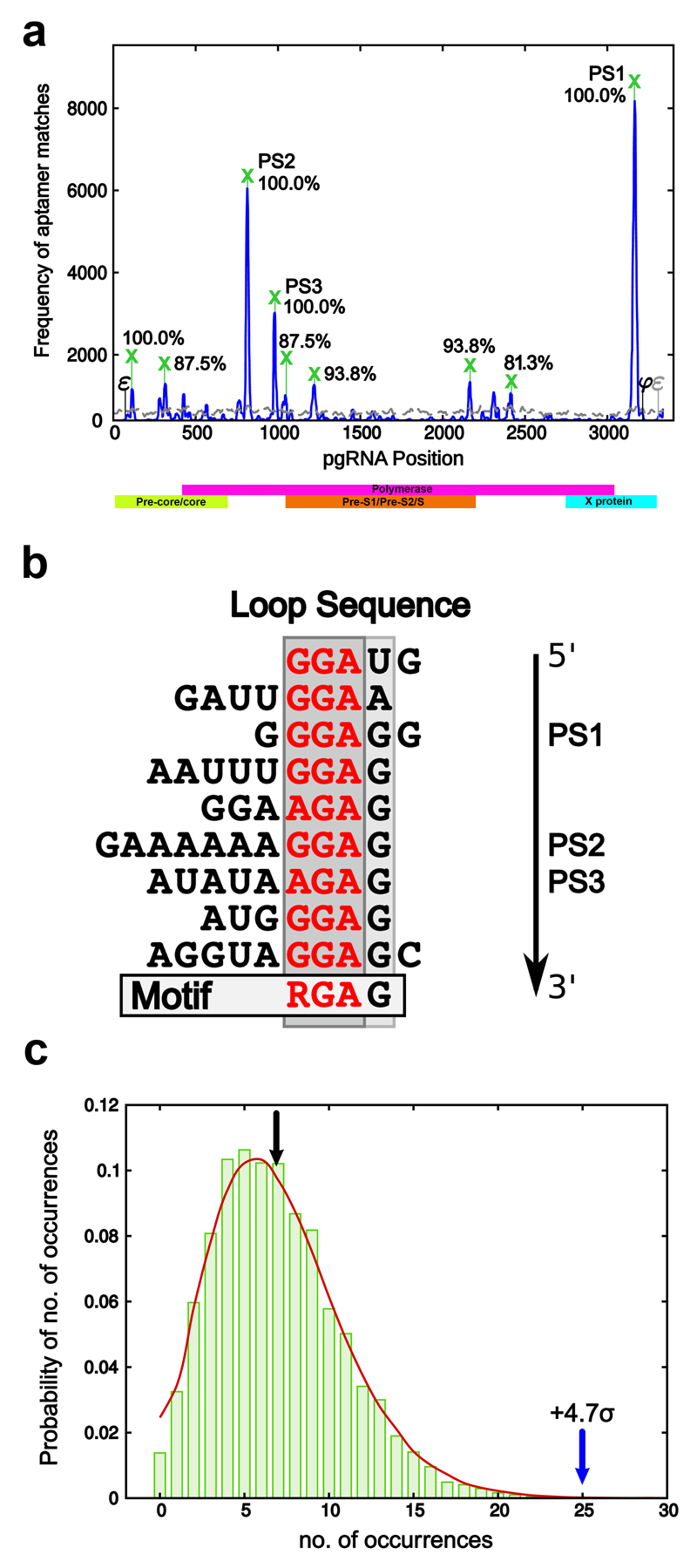
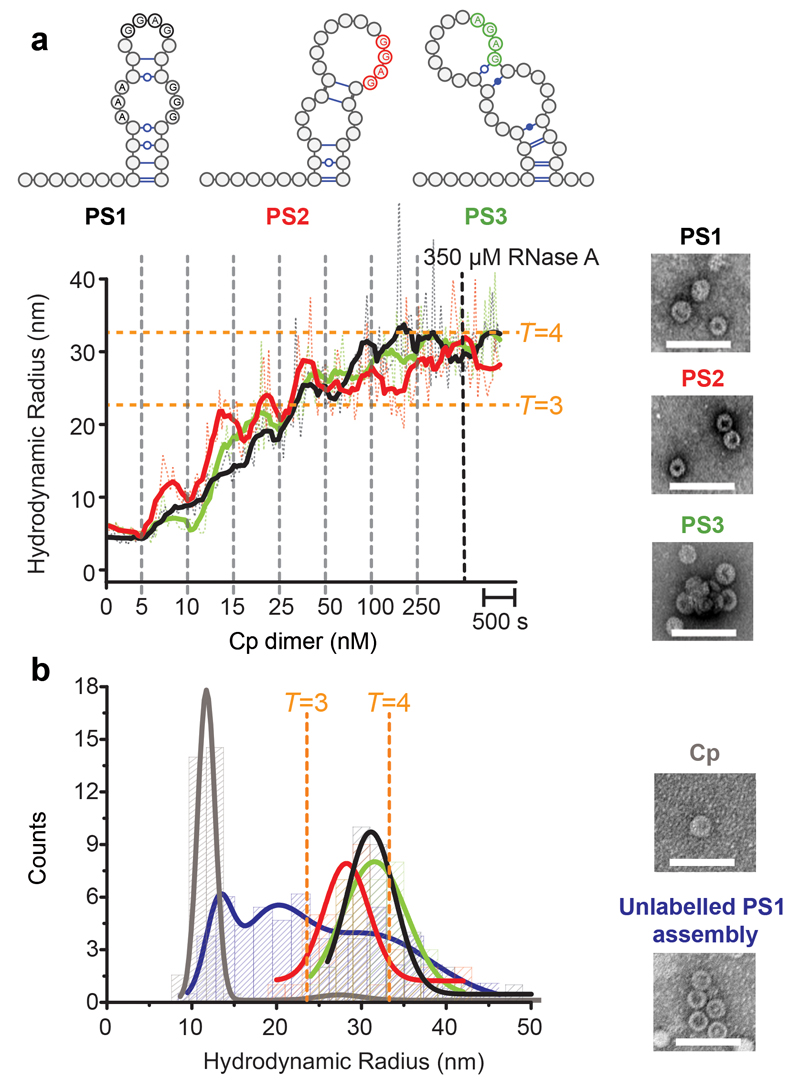
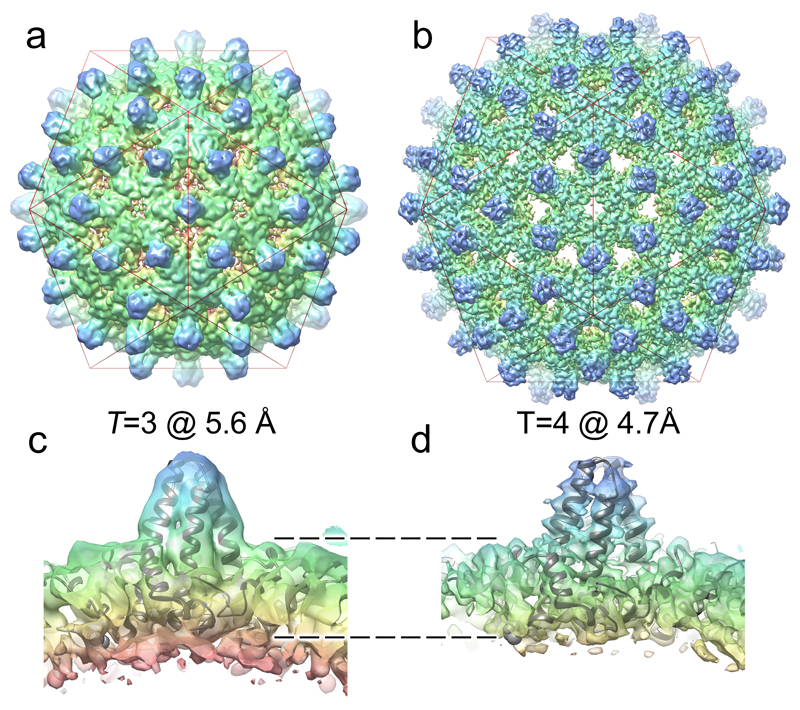
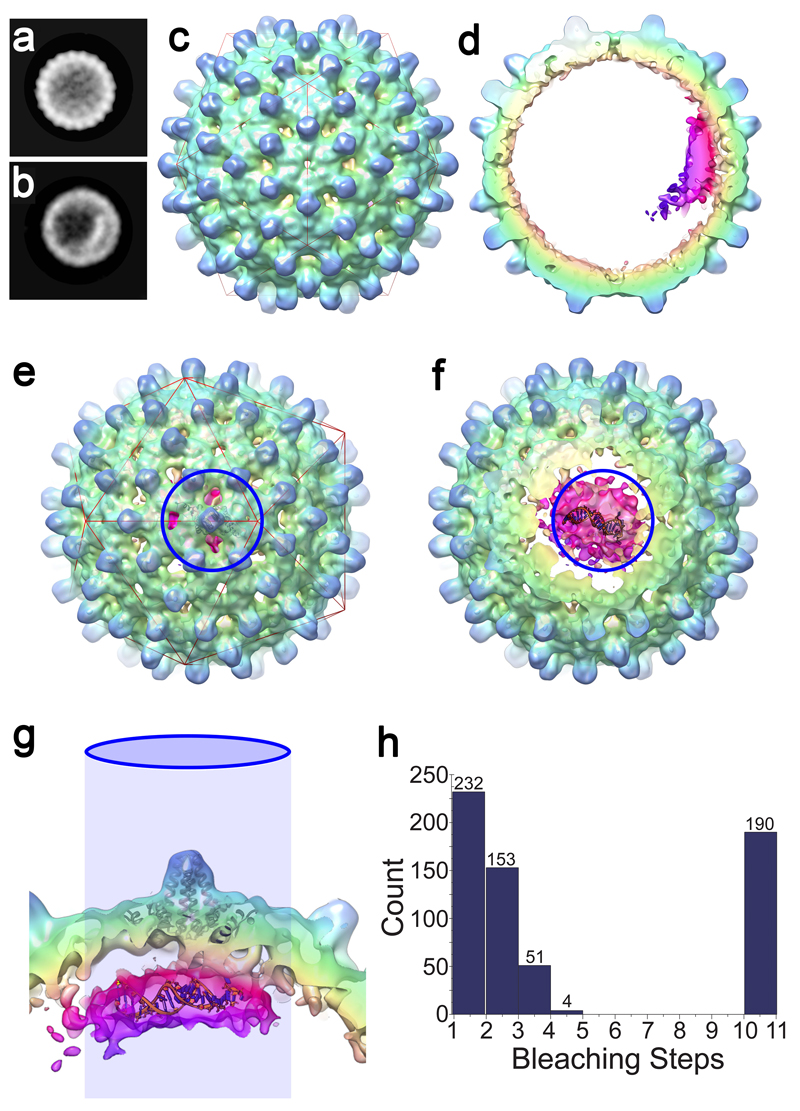
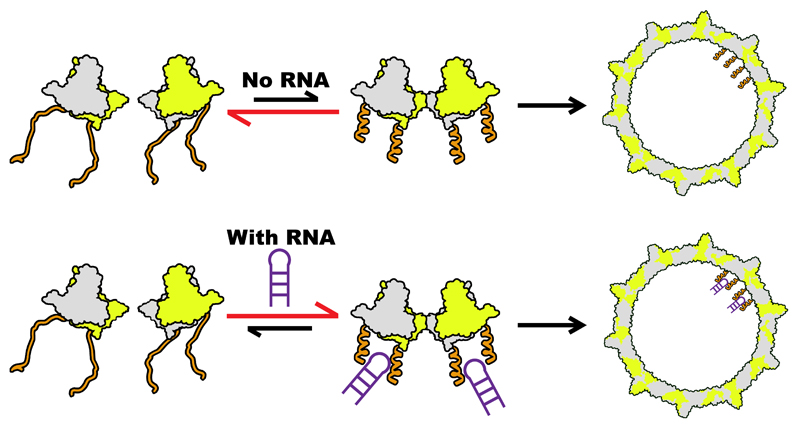
Formation of the hepatitis B virus nucleocapsid is an essential step in the viral lifecycle, but its assembly is not fully understood. We report the discovery of sequence-specific interactions between the viral pre-genome and the hepatitis B core protein that play roles in defining the nucleocapsid assembly pathway. Using RNA SELEX and bioinformatics, we identified multiple regions in the pre-genomic RNA with high affinity for core protein dimers. These RNAs form stem-loops with a conserved loop motif that trigger sequence-specific assembly of virus-like particles (VLPs) at much higher fidelity and yield than in the absence of RNA. The RNA oligos do not interact with preformed RNA-free VLPs, so their effects must occur during particle assembly. Asymmetric cryo-electron microscopy reconstruction of the T = 4 VLPs assembled in the presence of one of the RNAs reveals a unique internal feature connected to the main core protein shell via lobes of density. Biophysical assays suggest that this is a complex involving several RNA oligos interacting with the C-terminal arginine-rich domains of core protein. These core protein-RNA contacts may play one or more roles in regulating the organization of the pre-genome during nucleocapsid assembly, facilitating subsequent reverse transcription and acting as a nucleation complex for nucleocapsid assembly.
Conflict of interest statement
AZ is a co-founder and consultant of Assembly BioSciences.
Research in the Zlotnick lab is supported by the NIH and Assembly.
No Assembly BioSciences employee contributed to Dr. Zlotnick’s contribution to this work.
Figures
Comment in
-
Viral infection: Packing to leave.Nat Rev Microbiol. 2017 Aug;15(8):450-451. doi: 10.1038/nrmicro.2017.81. Epub 2017 Jul 10. Nat Rev Microbiol. 2017. PMID: 28690322 No abstract available.
References
-
- WHO. Weekly epidemiological record. WHO. 2009;84:405–420.
-
- Murray K, et al. Protective immunisation against hepatitis B with an internal antigen of the virus. J Med Virol. 1987;23:101–107. - PubMed
-
- Nassal M. Hepatitis B viruses: reverse transcription a different way. Virus Res. 2008;134:235–249. - PubMed
-
- Seeger C, Zoulim F, Mason WS. Hepadnaviruses. Vol. 2. Fields Virology; 2007. pp. 2977–3209.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
