

The glutamate/cystine antiporter SLC7A11/xCT enhances cancer cell dependency on glucose by exporting glutamate
- PMID: 28630042
- PMCID: PMC5572906
- DOI: 10.1074/jbc.M117.798405
The glutamate/cystine antiporter SLC7A11/xCT enhances cancer cell dependency on glucose by exporting glutamate
Abstract
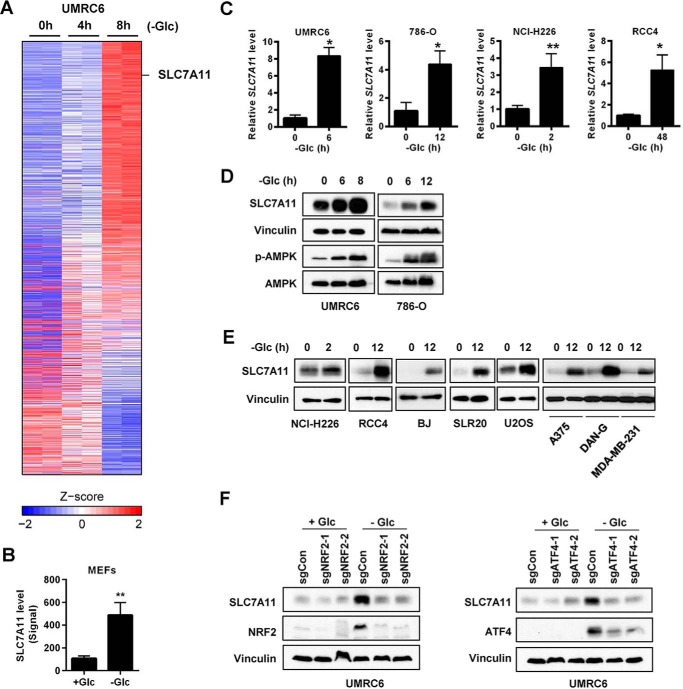
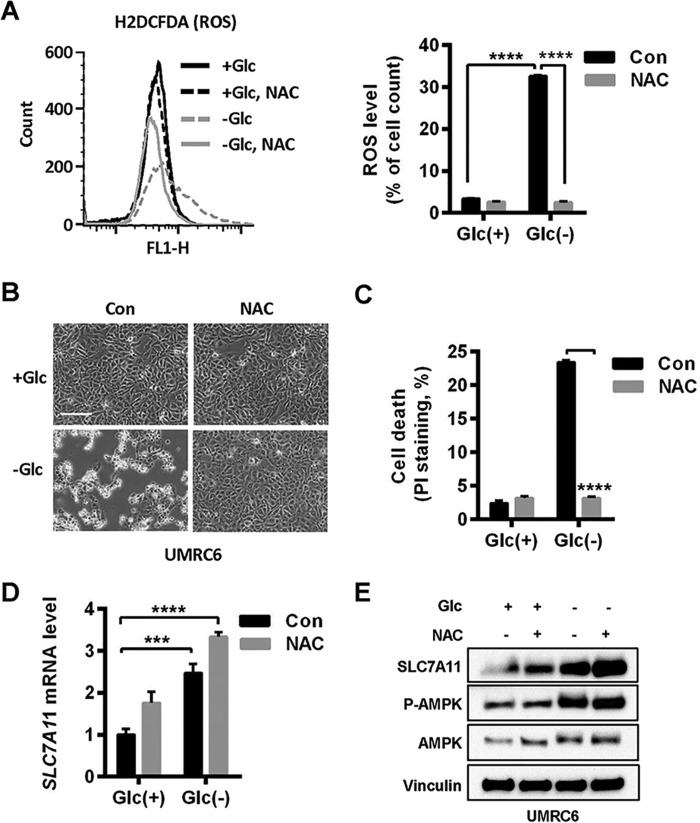
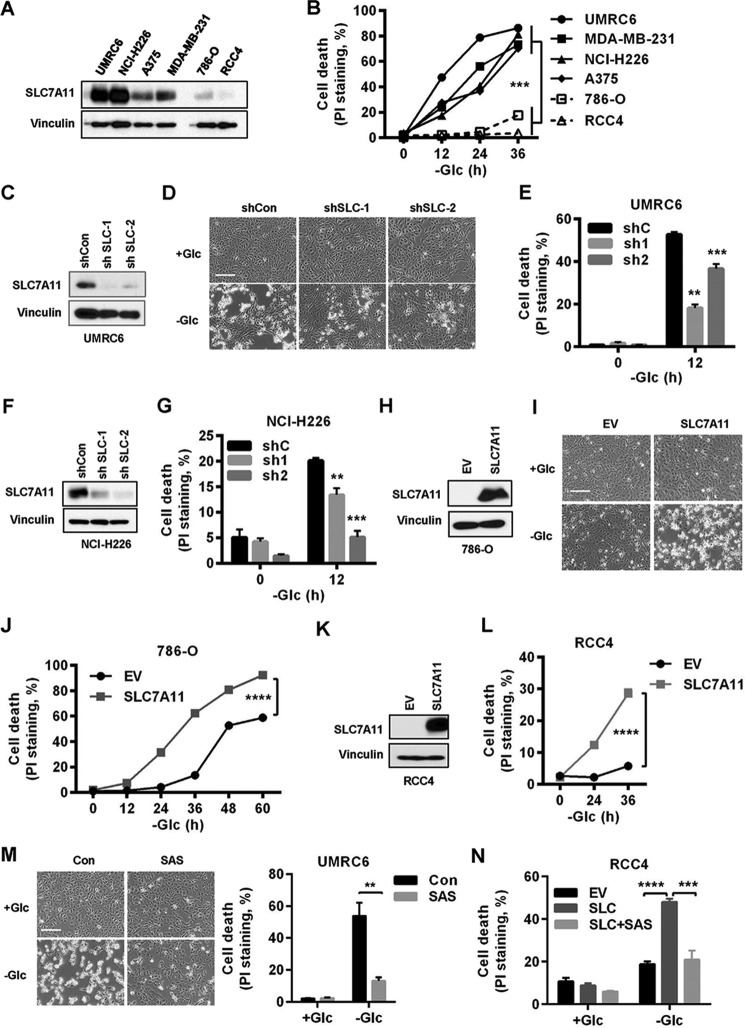
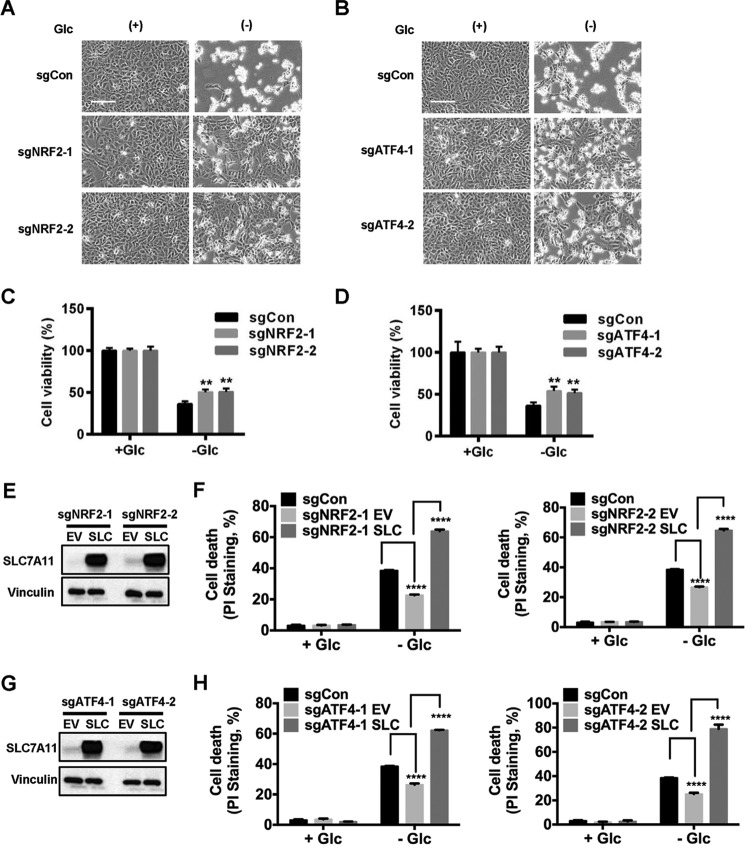
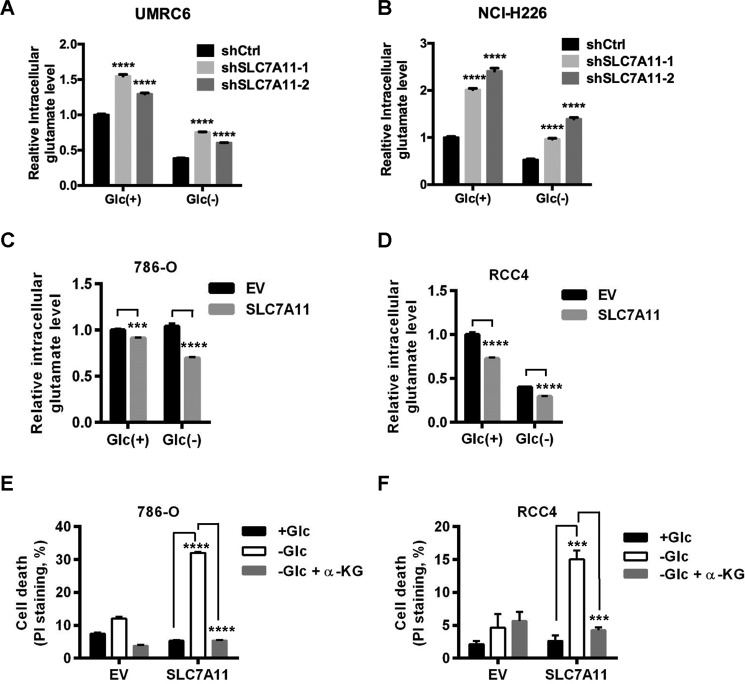
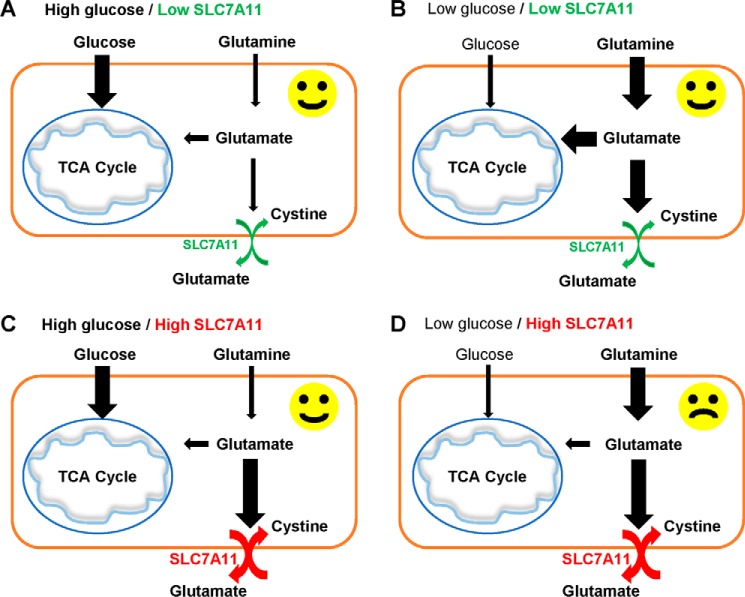
Cancer cells with specific genetic alterations may be highly dependent on certain nutrients for survival, which can inform therapeutic strategies to target these cancer-specific metabolic vulnerabilities. The glutamate/cystine antiporter solute carrier family 7 member 11 (SLC7A11, also called xCT) is overexpressed in several cancers. Contrasting the established pro-survival roles of SLC7A11 under other stress conditions, here we report the unexpected finding that SLC7A11 overexpression enhances cancer cell dependence on glucose and renders cancer cells more sensitive to glucose starvation-induced cell death and, conversely, that SLC7A11 deficiency by either knockdown or pharmacological inhibition promotes cancer cell survival upon glucose starvation. We further show that glucose starvation induces SLC7A11 expression through ATF4 and NRF2 transcription factors and, correspondingly, that ATF4 or NRF2 deficiency also renders cancer cells more resistant to glucose starvation. Finally, we show that SLC7A11 overexpression decreases whereas SLC7A11 deficiency increases intracellular glutamate levels because of SLC7A11-mediated glutamate export and that supplementation of α-ketoglutarate, a key downstream metabolite of glutamate, fully restores survival in SLC7A11-overexpressing cells under glucose starvation. Together, our results support the notion that both glucose and glutamate have important roles in maintaining cancer cell survival and uncover a previously unappreciated role of SLC7A11 to promote cancer cell dependence on glucose. Our study therefore informs therapeutic strategies to target the metabolic vulnerability in tumors with high SLC7A11 expression.
Keywords: SLC7A11; cell death; glucose; glutamate; reactive oxygen species (ROS); tumor metabolism; xCT.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
