

Rapid Evolution of Ovarian-Biased Genes in the Yellow Fever Mosquito (Aedes aegypti)
- PMID: 28630112
- PMCID: PMC5560810
- DOI: 10.1534/genetics.117.201343
Rapid Evolution of Ovarian-Biased Genes in the Yellow Fever Mosquito (Aedes aegypti)
Abstract
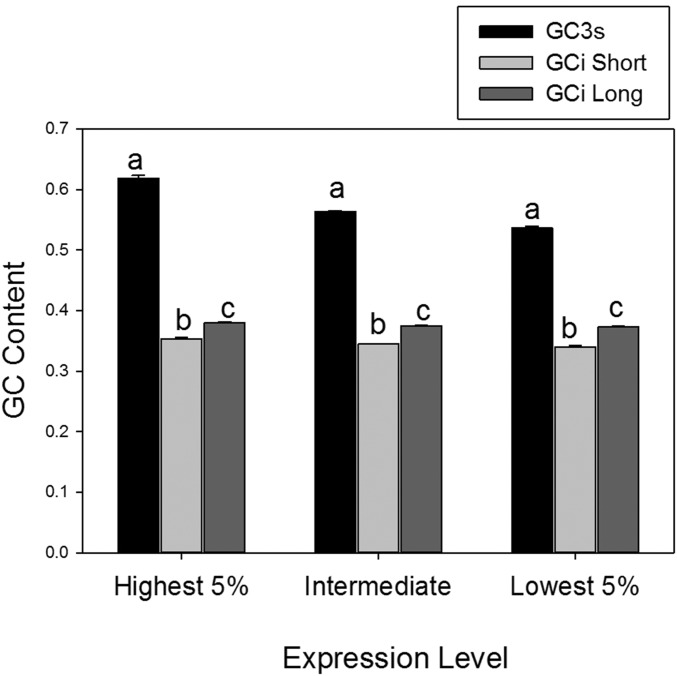
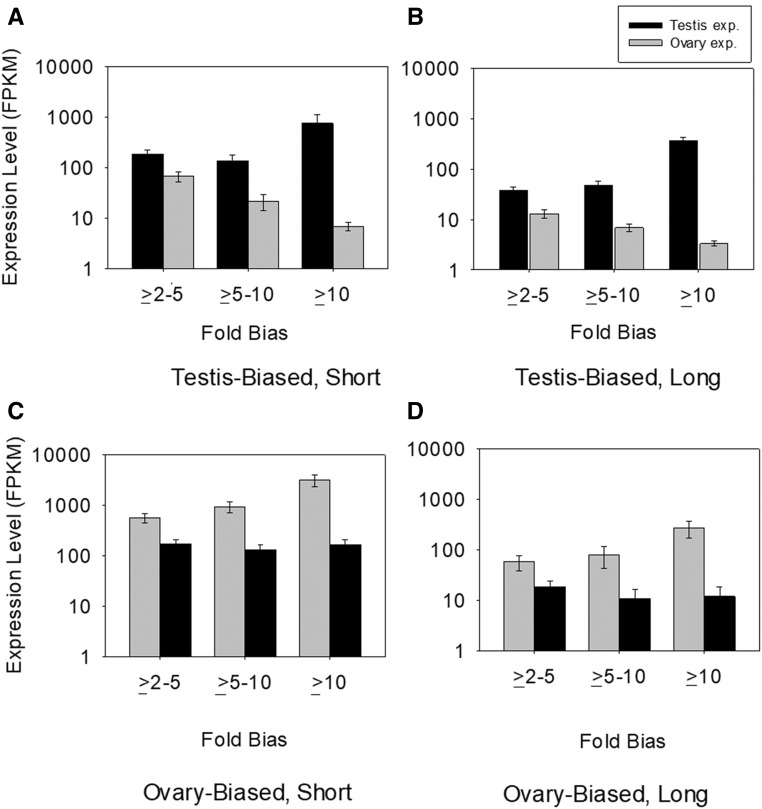
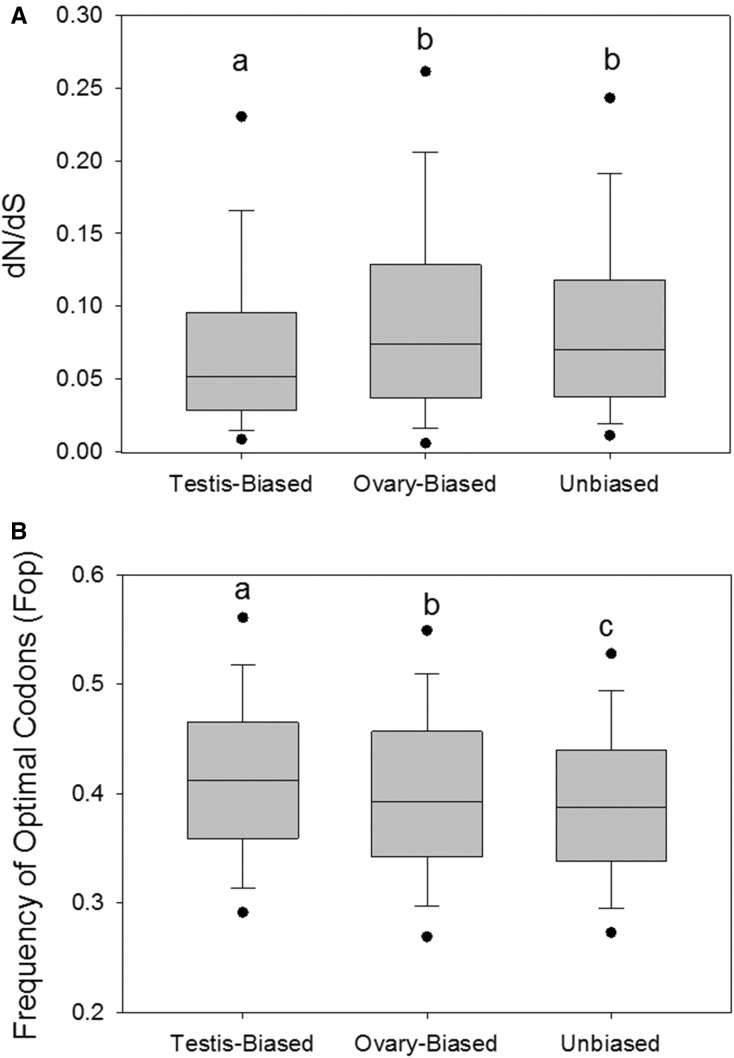
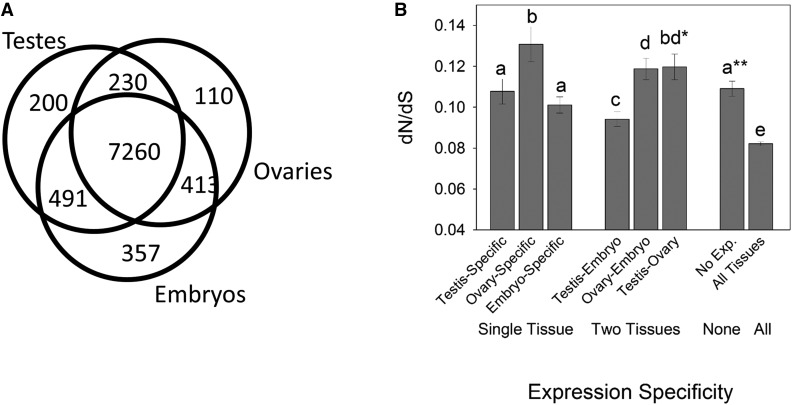
Males and females exhibit highly dimorphic phenotypes, particularly in their gonads, which is believed to be driven largely by differential gene expression. Typically, the protein sequences of genes upregulated in males, or male-biased genes, evolve rapidly as compared to female-biased and unbiased genes. To date, the specific study of gonad-biased genes remains uncommon in metazoans. Here, we identified and studied a total of 2927, 2013, and 4449 coding sequences (CDS) with ovary-biased, testis-biased, and unbiased expression, respectively, in the yellow fever mosquito Aedes aegypti The results showed that ovary-biased and unbiased CDS had higher nonsynonymous to synonymous substitution rates (dN/dS) and lower optimal codon usage (those codons that promote efficient translation) than testis-biased genes. Further, we observed higher dN/dS in ovary-biased genes than in testis-biased genes, even for genes coexpressed in nonsexual (embryo) tissues. Ovary-specific genes evolved exceptionally fast, as compared to testis- or embryo-specific genes, and exhibited higher frequency of positive selection. Genes with ovary expression were preferentially involved in olfactory binding and reception. We hypothesize that at least two potential mechanisms could explain rapid evolution of ovary-biased genes in this mosquito: (1) the evolutionary rate of ovary-biased genes may be accelerated by sexual selection (including female-female competition or male-mate choice) affecting olfactory genes during female swarming by males, and/or by adaptive evolution of olfactory signaling within the female reproductive system (e.g., sperm-ovary signaling); and/or (2) testis-biased genes may exhibit decelerated evolutionary rates due to the formation of mating plugs in the female after copulation, which limits male-male sperm competition.
Keywords: Aedes aegypti; Genetics of Sex; gonads; molecular evolution; ovary-biased genes; protein sequence divergence.
Copyright © 2017 by the Genetics Society of America.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
