

Toward Universal Forward Genetics: Using a Draft Genome Sequence of the Nematode Oscheius tipulae To Identify Mutations Affecting Vulva Development
- PMID: 28630114
- PMCID: PMC5560785
- DOI: 10.1534/genetics.117.203521
Toward Universal Forward Genetics: Using a Draft Genome Sequence of the Nematode Oscheius tipulae To Identify Mutations Affecting Vulva Development
Abstract
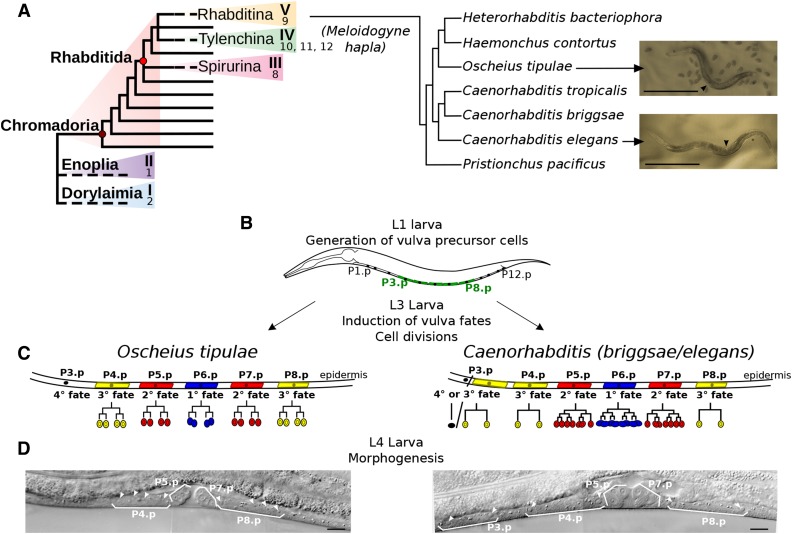
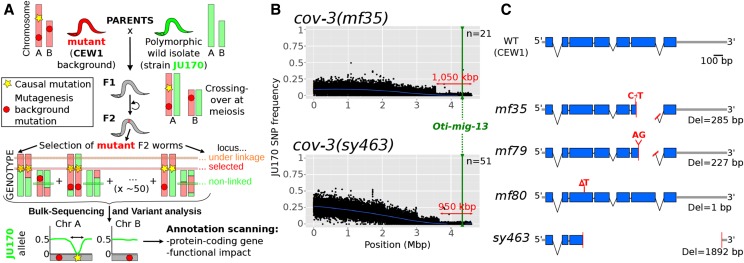
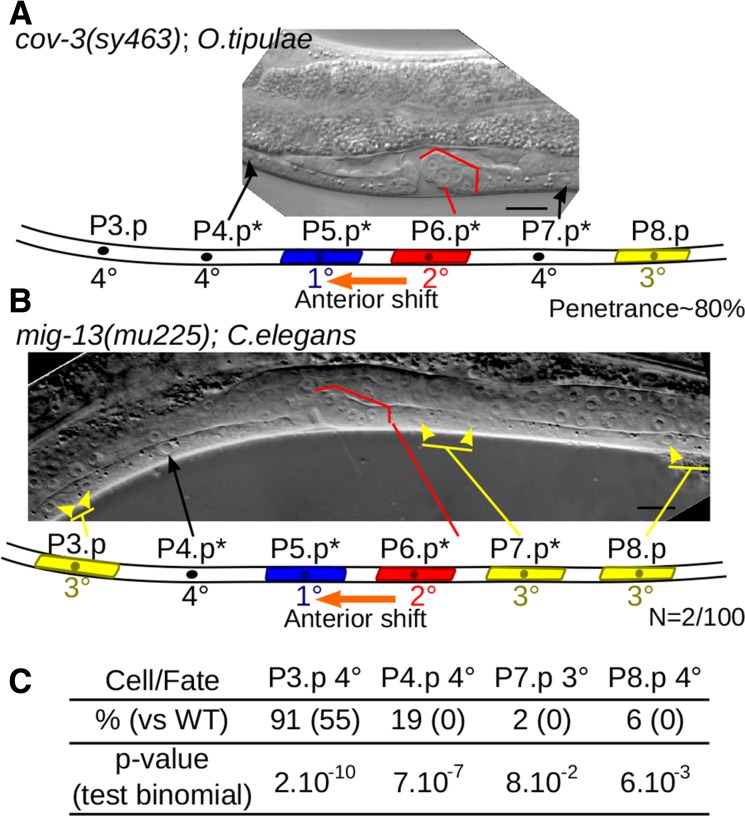
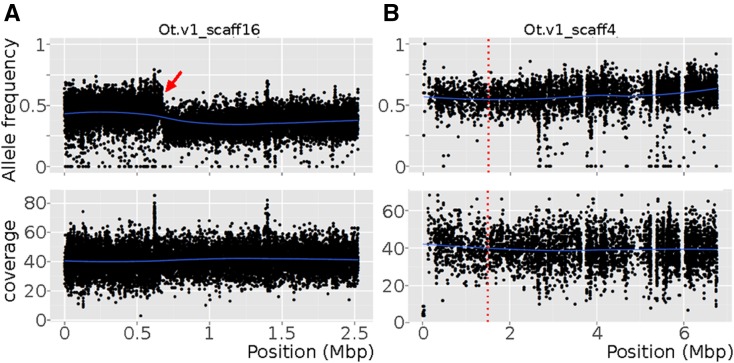
Mapping-by-sequencing has become a standard method to map and identify phenotype-causing mutations in model species. Here, we show that a fragmented draft assembly is sufficient to perform mapping-by-sequencing in nonmodel species. We generated a draft assembly and annotation of the genome of the free-living nematode Oscheius tipulae, a distant relative of the model Caenorhabditis elegans We used this draft to identify the likely causative mutations at the O. tipulae cov-3 locus, which affect vulval development. The cov-3 locus encodes the O. tipulae ortholog of C. elegans mig-13, and we further show that Cel-mig-13 mutants also have an unsuspected vulval-development phenotype. In a virtuous circle, we were able to use the linkage information collected during mutant mapping to improve the genome assembly. These results showcase the promise of genome-enabled forward genetics in nonmodel species.
Keywords: Oscheius tipulae; genome assembly; mapping-by-sequencing; mig-13; vulva development.
Copyright © 2017 by the Genetics Society of America.
Figures
References
-
- Abe A., Kosugi S., Yoshida K., Natsume S., Takagi H., et al. , 2012. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 30: 174–178. - PubMed
-
- Ahn I. Y., Winter C. E., 2006. The genome of Oscheius tipulae: determination of size, complexity, and structure by DNA reassociation using fluorescent dye. Genome 49: 1007–1015. - PubMed
-
- Anderson P., 1995. Mutagenesis. Methods Cell Biol. 48: 31–58. - PubMed
-
- Andrews, S., 2010 FastQC A Quality Control Tool for High Throughput Sequence Data. http://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
-
- Baïlle D., Barrière A., Félix M.-A., 2008. Oscheius tipulae, a widespread hermaphroditic soil nematode, displays a higher genetic diversity and geographical structure than Caenorhabditis elegans. Mol. Ecol. 17: 1523–1534. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
