

The Small GTPase Rac1 Contributes to Extinction of Aversive Memories of Drug Withdrawal by Facilitating GABAA Receptor Endocytosis in the vmPFC
- PMID: 28630256
- PMCID: PMC6705728
- DOI: 10.1523/JNEUROSCI.3859-16.2017
The Small GTPase Rac1 Contributes to Extinction of Aversive Memories of Drug Withdrawal by Facilitating GABAA Receptor Endocytosis in the vmPFC
Abstract
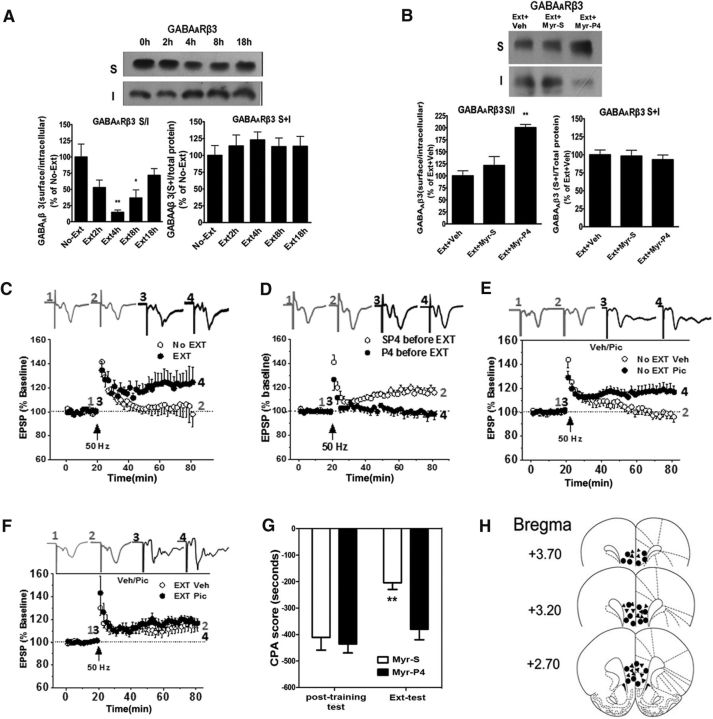
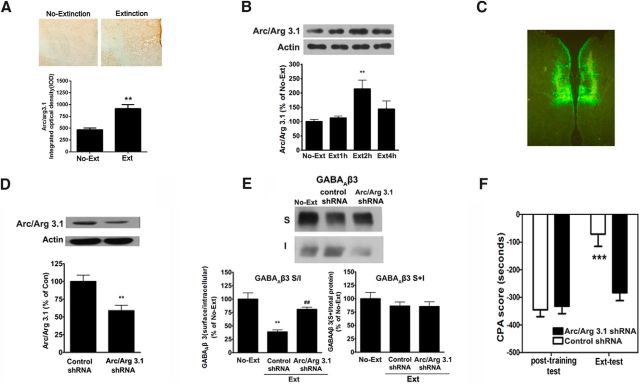
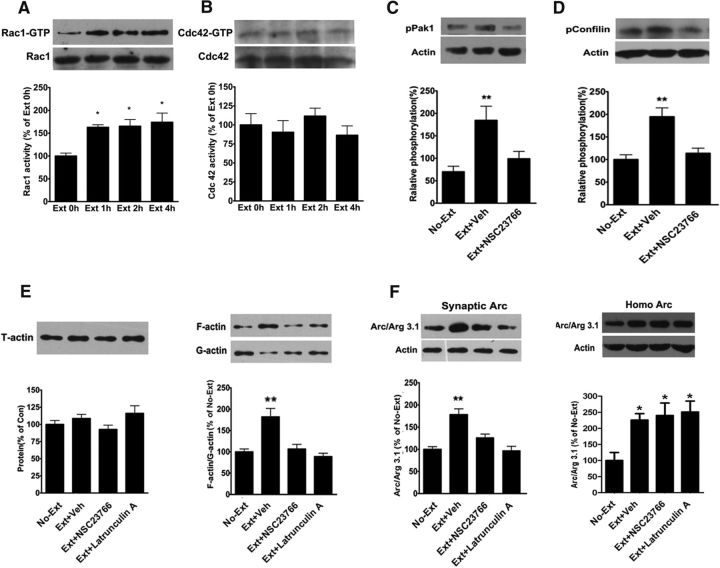
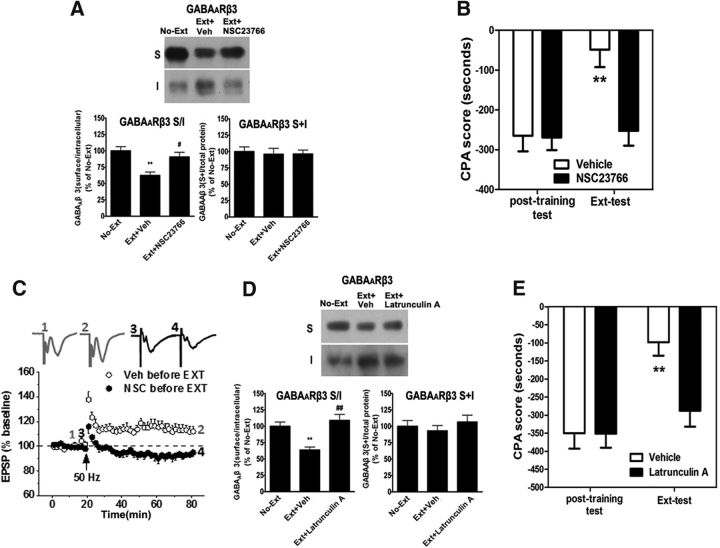
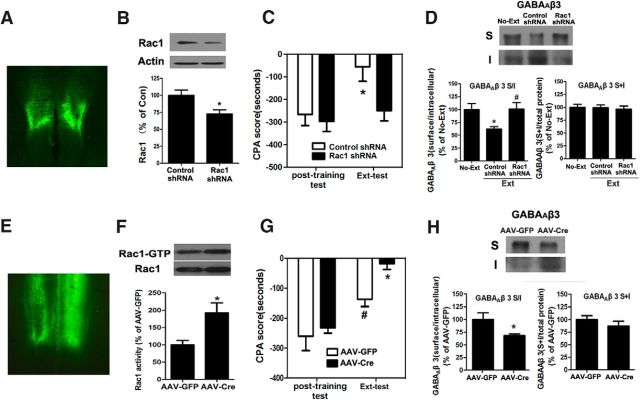
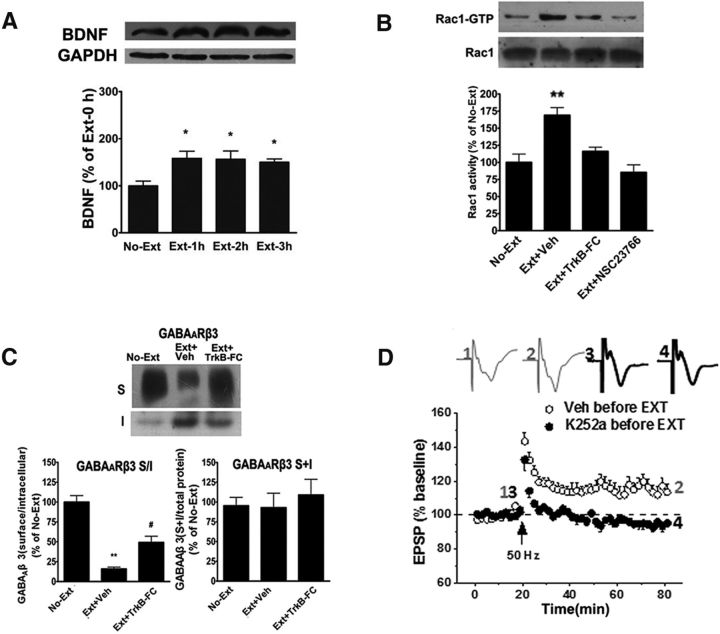
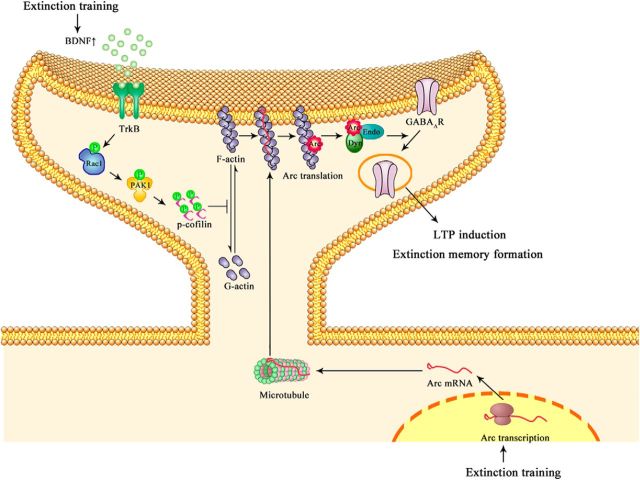
Extinction of aversive memories has been a major concern in neuropsychiatric disorders, such as anxiety disorders and drug addiction. However, the mechanisms underlying extinction of aversive memories are not fully understood. Here, we report that extinction of conditioned place aversion (CPA) to naloxone-precipitated opiate withdrawal in male rats activates Rho GTPase Rac1 in the ventromedial prefrontal cortex (vmPFC) in a BDNF-dependent manner, which determines GABAA receptor (GABAAR) endocytosis via triggering synaptic translocation of activity-regulated cytoskeleton-associated protein (Arc) through facilitating actin polymerization. Active Rac1 is essential and sufficient for GABAAR endocytosis and CPA extinction. Knockdown of Rac1 expression within the vmPFC of rats using Rac1-shRNA suppressed GABAAR endocytosis and CPA extinction, whereas expression of a constitutively active form of Rac1 accelerated GABAAR endocytosis and CPA extinction. The crucial role of GABAAR endocytosis in the LTP induction and CPA extinction is evinced by the findings that blockade of GABAAR endocytosis by a dynamin function-blocking peptide (Myr-P4) abolishes LTP induction and CPA extinction. Thus, the present study provides first evidence that Rac1-dependent GABAAR endocytosis plays a crucial role in extinction of aversive memories and reveals the sequence of molecular events that contribute to learning experience modulation of synaptic GABAAR endocytosis.SIGNIFICANCE STATEMENT This study reveals that Rac1-dependent GABAAR endocytosis plays a crucial role in extinction of aversive memories associated with drug withdrawal and identifies Arc as a downstream effector of Rac1 regulations of synaptic plasticity as well as learning and memory, thereby suggesting therapeutic targets to promote extinction of the unwanted memories.
Keywords: GABAA receptor; Rac1; aversive memory; drug withdrawal; endocytosis; extinction.
Copyright © 2017 the authors 0270-6474/17/377096-15$15.00/0.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials