

Bacteria-mediated hypoxia functions as a signal for mosquito development
- PMID: 28630299
- PMCID: PMC5502624
- DOI: 10.1073/pnas.1702983114
Bacteria-mediated hypoxia functions as a signal for mosquito development
Abstract
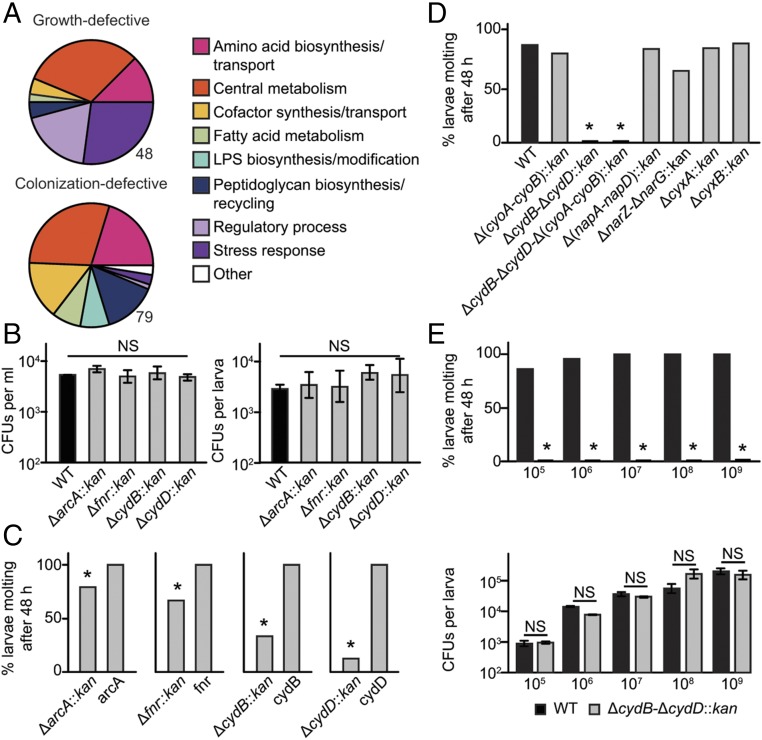
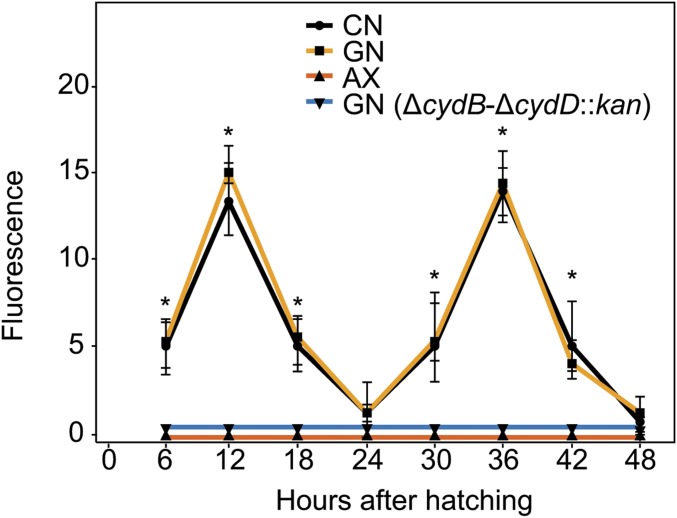
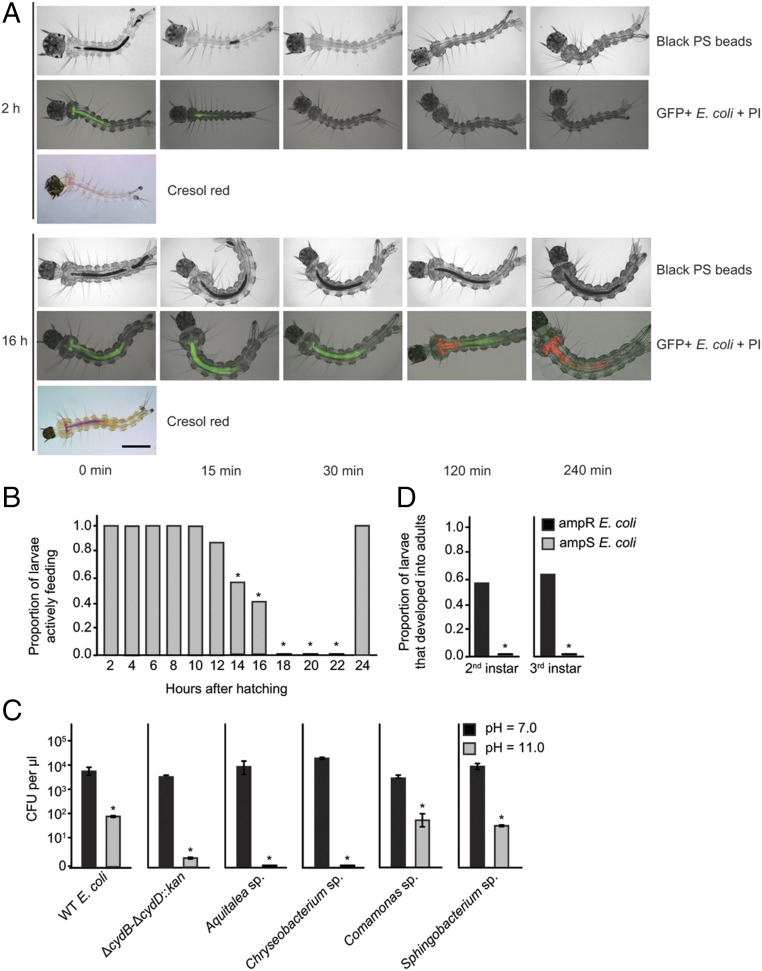
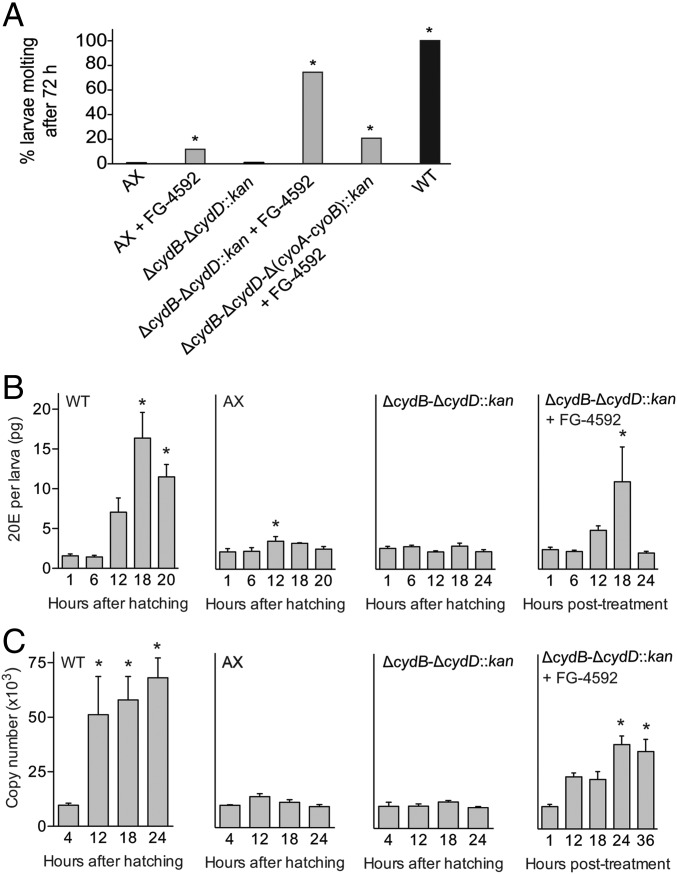
Mosquitoes host communities of microbes in their digestive tract that consist primarily of bacteria. We previously reported that several mosquito species, including Aedes aegypti, do not develop beyond the first instar when fed a nutritionally complete diet in the absence of a gut microbiota. In contrast, several species of bacteria, including Escherichia coli, rescue development of axenic larvae into adults. The molecular mechanisms underlying bacteria-dependent growth are unknown. Here, we designed a genetic screen around E. coli that identified high-affinity cytochrome bd oxidase as an essential bacterial gene product for mosquito growth. Bioassays showed that bacteria in nonsterile larvae and gnotobiotic larvae inoculated with wild-type E. coli reduced midgut oxygen levels below 5%, whereas larvae inoculated with E. coli mutants defective for cytochrome bd oxidase did not. Experiments further supported that hypoxia leads to growth and ecdysone-induced molting. Altogether, our results identify aerobic respiration by bacteria as a previously unknown but essential process for mosquito development.
Keywords: bacteria; growth; hypoxia; insect; microbiota.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Engel P, Moran NA. The gut microbiota of insects–Diversity in structure and function. FEMS Microbiol Rev. 2013;37:699–735. - PubMed
-
- Sommer F, Bäckhed F. The gut microbiota–Masters of host development and physiology. Nat Rev Microbiol. 2013;11:227–238. - PubMed
-
- Merritt RW, Dadd RH, Walker ED. Feeding behavior, natural food, and nutritional relationships of larval mosquitoes. Annu Rev Entomol. 1992;37:349–376. - PubMed
-
- Clements AN. The Biology of Mosquitoes, Development, Nutrition, and Reproduction. Vol 1 Chapman & Hall; New York: 1992.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
