

TCTE1 is a conserved component of the dynein regulatory complex and is required for motility and metabolism in mouse spermatozoa
- PMID: 28630322
- PMCID: PMC5502601
- DOI: 10.1073/pnas.1621279114
TCTE1 is a conserved component of the dynein regulatory complex and is required for motility and metabolism in mouse spermatozoa
Abstract
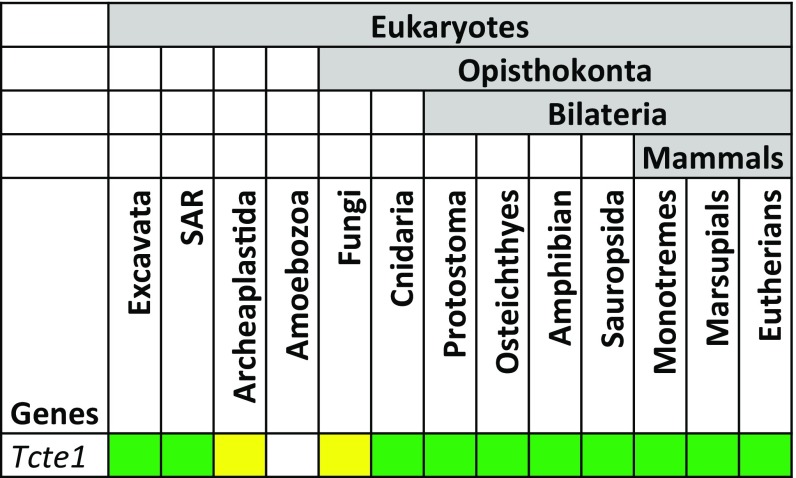
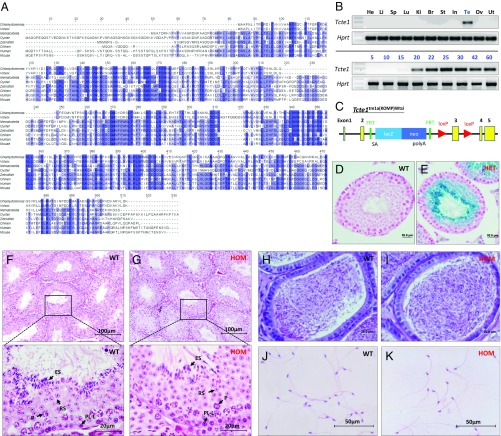
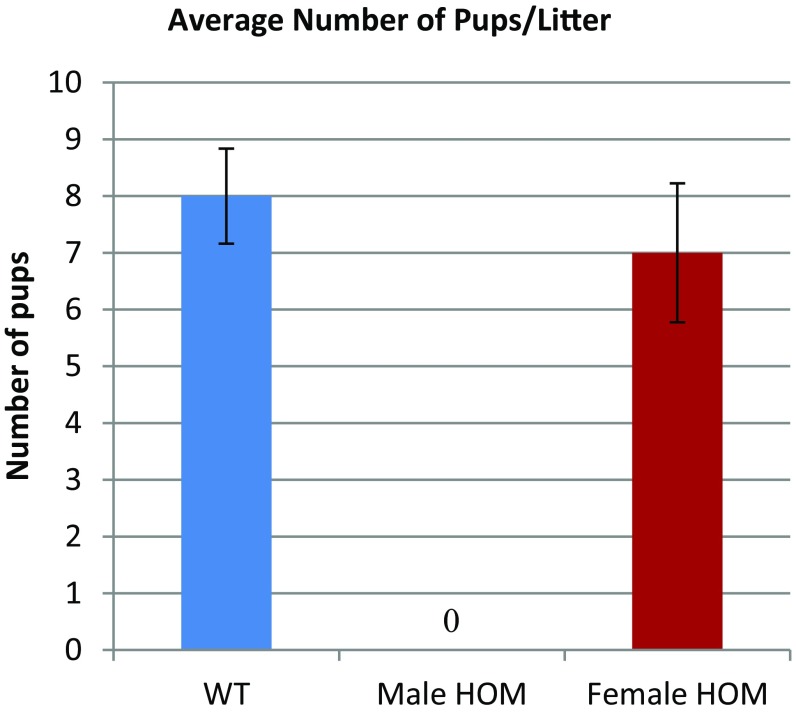
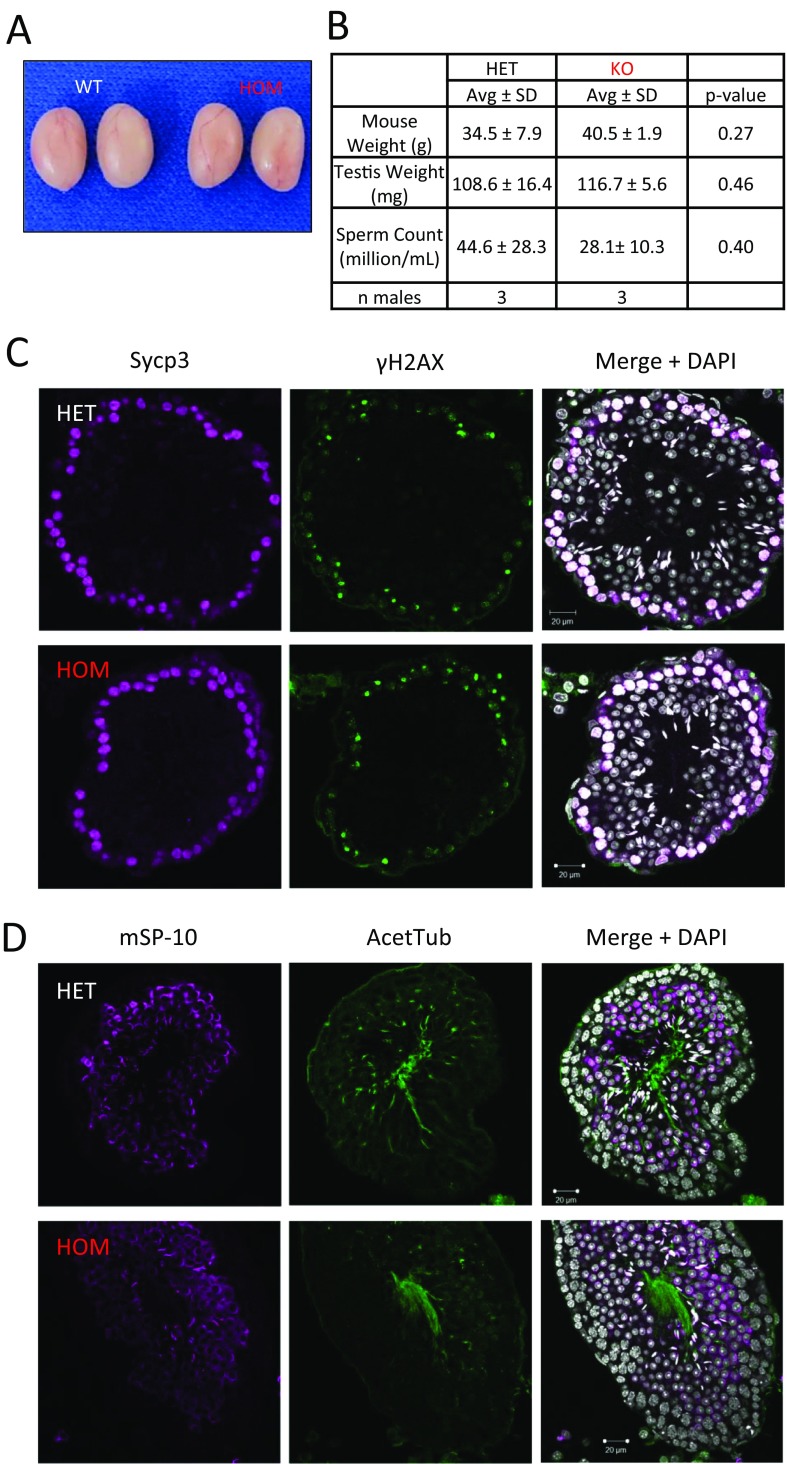
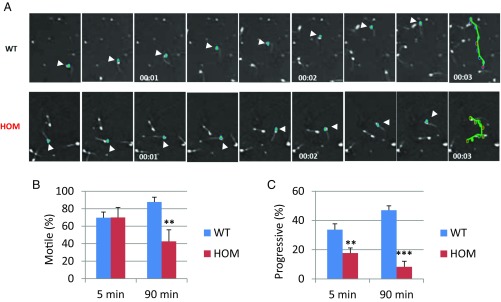
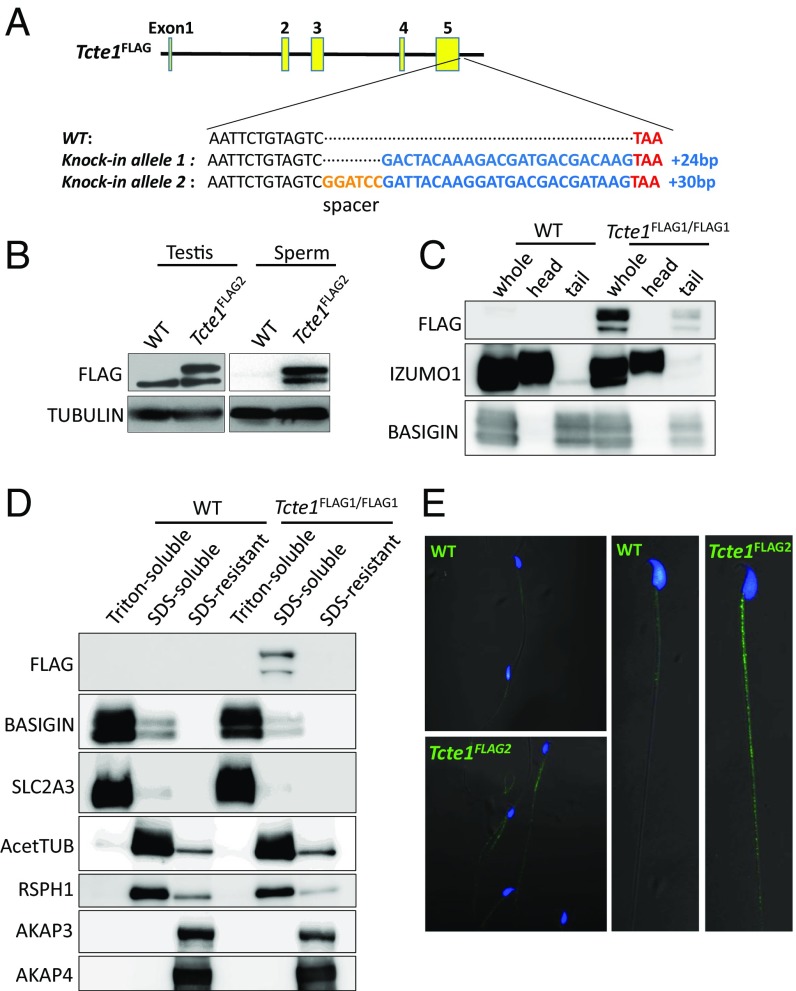
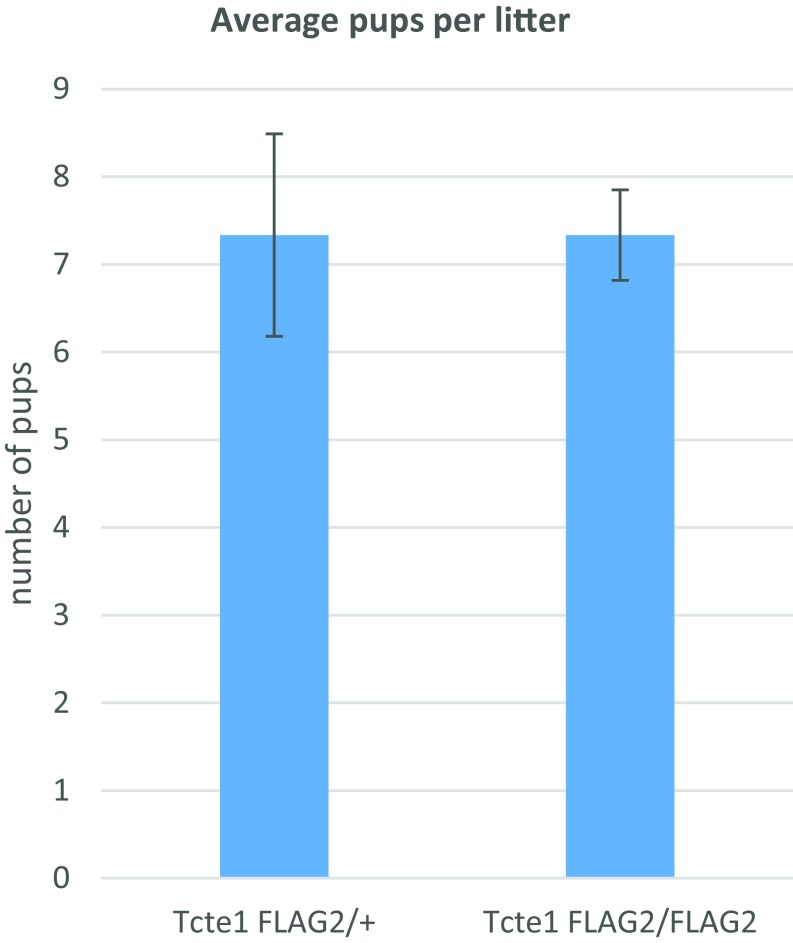
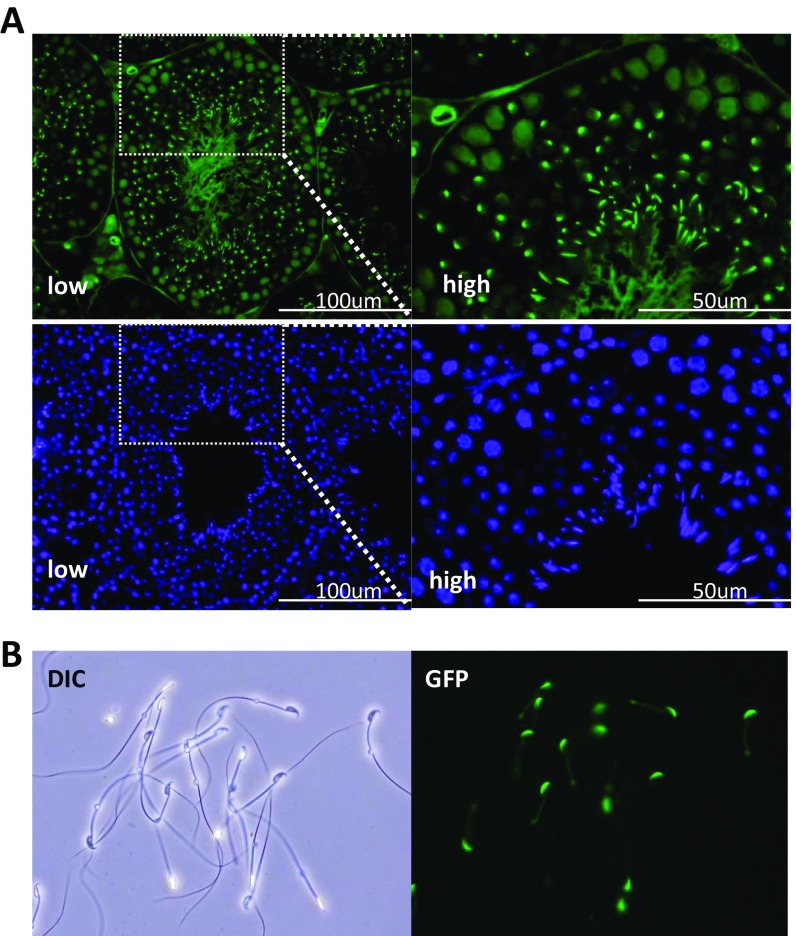
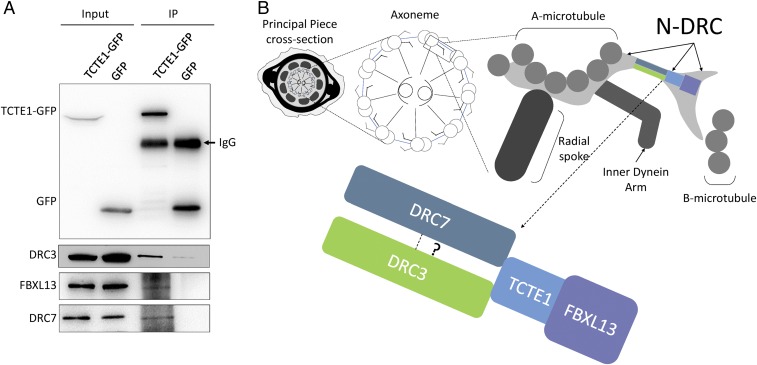
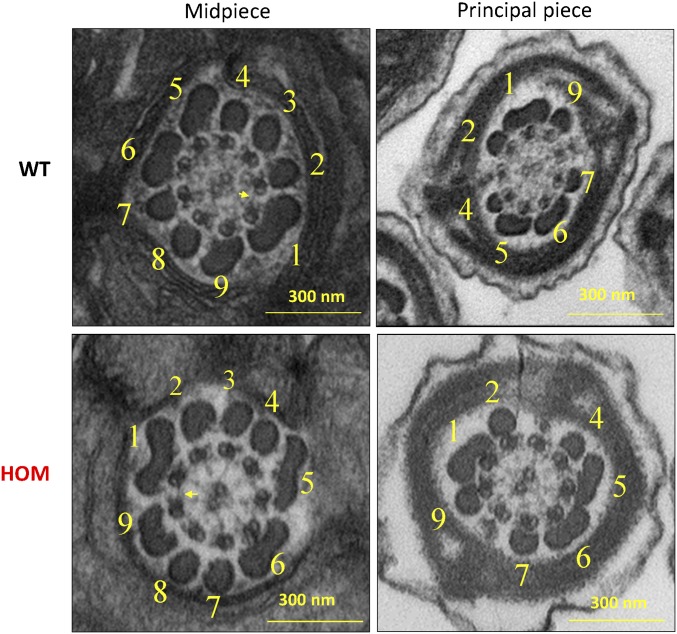
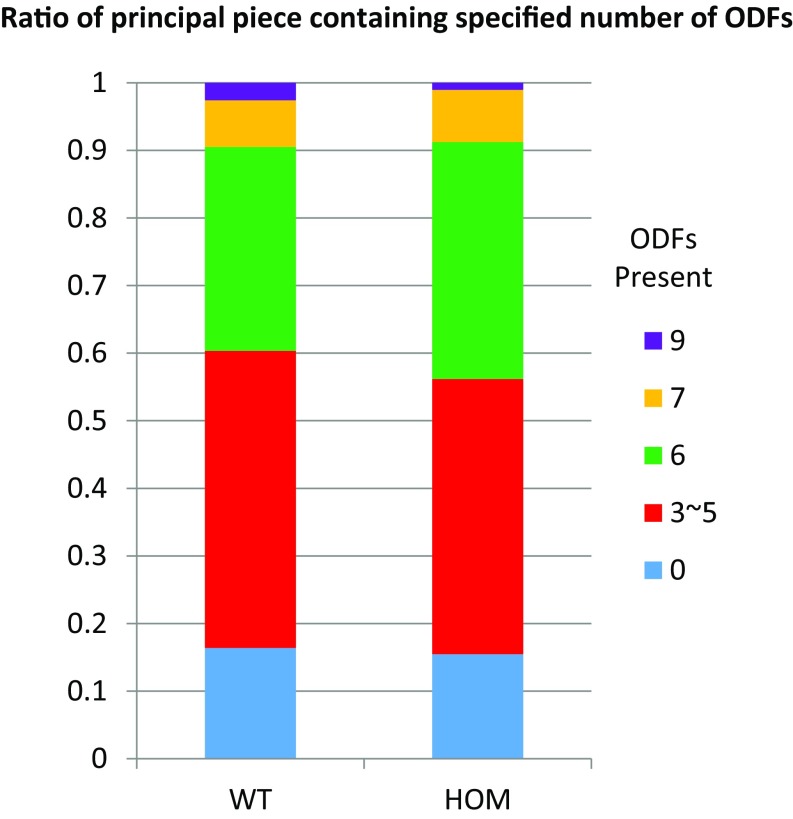
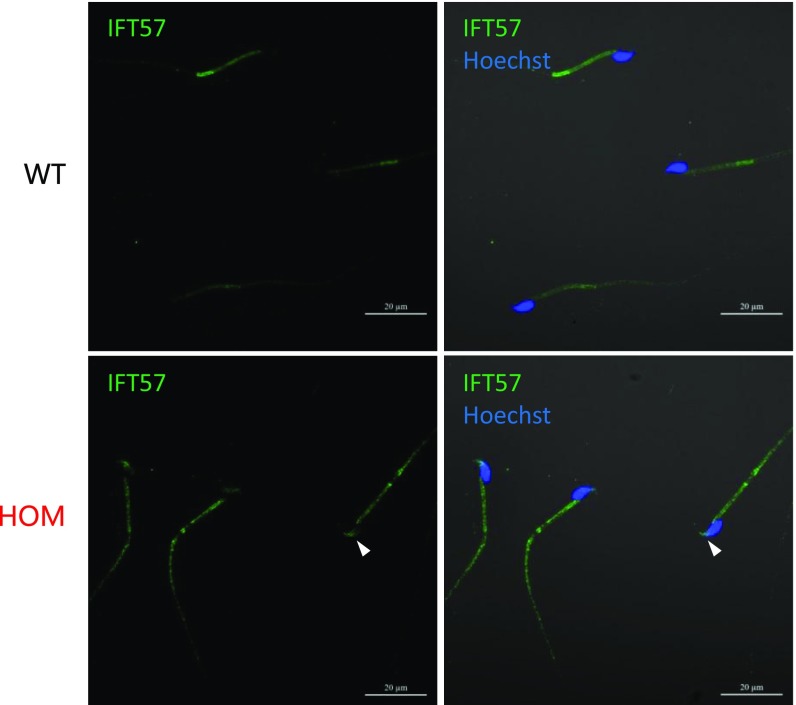
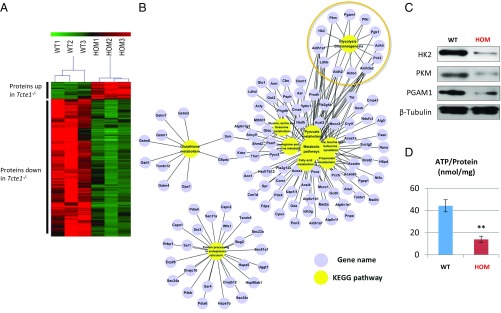
Flagella and cilia are critical cellular organelles that provide a means for cells to sense and progress through their environment. The central component of flagella and cilia is the axoneme, which comprises the "9+2" microtubule arrangement, dynein arms, radial spokes, and the nexin-dynein regulatory complex (N-DRC). Failure to properly assemble components of the axoneme leads to defective flagella and in humans leads to a collection of diseases referred to as ciliopathies. Ciliopathies can manifest as severe syndromic diseases that affect lung and kidney function, central nervous system development, bone formation, visceral organ organization, and reproduction. T-Complex-Associated-Testis-Expressed 1 (TCTE1) is an evolutionarily conserved axonemal protein present from Chlamydomonas (DRC5) to mammals that localizes to the N-DRC. Here, we show that mouse TCTE1 is testis-enriched in its expression, with its mRNA appearing in early round spermatids and protein localized to the flagellum. TCTE1 is 498 aa in length with a leucine rich repeat domain at the C terminus and is present in eukaryotes containing a flagellum. Knockout of Tcte1 results in male sterility because Tcte1-null spermatozoa show aberrant motility. Although the axoneme is structurally normal in Tcte1 mutant spermatozoa, Tcte1-null sperm demonstrate a significant decrease of ATP, which is used by dynein motors to generate the bending force of the flagellum. These data provide a link to defining the molecular intricacies required for axoneme function, sperm motility, and male fertility.
Keywords: asthenozoospermia; glycolysis; male infertility; mutant mouse; testis-specific gene.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Faguy DM, Jarrell KF, Kuzio J, Kalmokoff ML. Molecular analysis of archael flagellins: Similarity to the type IV pilin-transport superfamily widespread in bacteria. Can J Microbiol. 1994;40:67–71. - PubMed
-
- Lindemann CB, Lesich KA. Functional anatomy of the mammalian sperm flagellum. Cytoskeleton. 2016;73:652–669. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
