

The case for defined protein folding pathways
- PMID: 28630329
- PMCID: PMC5547639
- DOI: 10.1073/pnas.1706196114
The case for defined protein folding pathways
Abstract
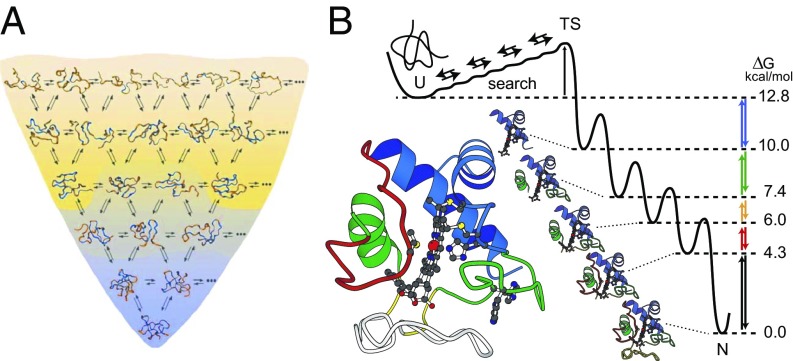
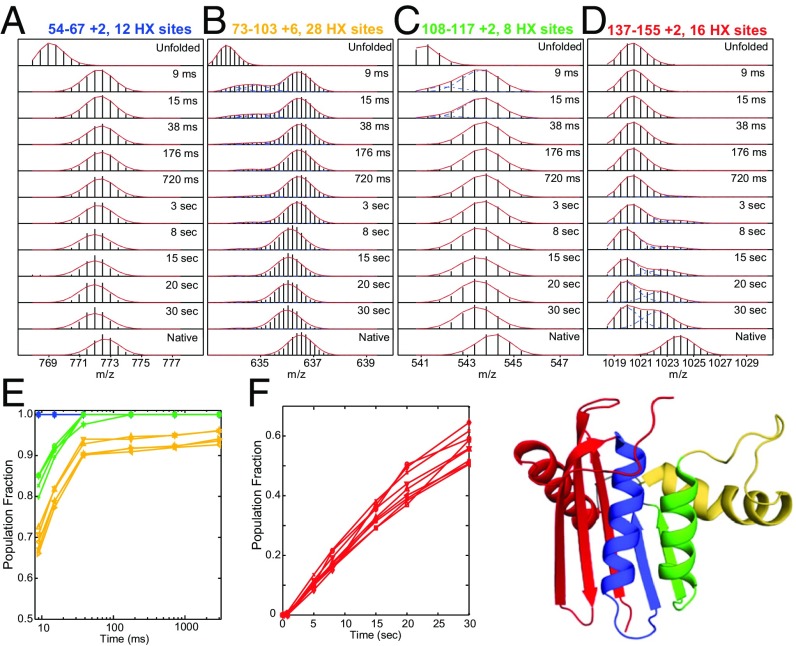
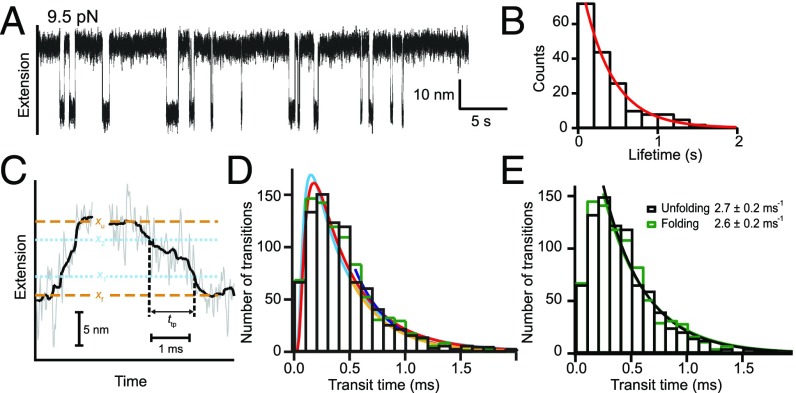
We consider the differences between the many-pathway protein folding model derived from theoretical energy landscape considerations and the defined-pathway model derived from experiment. A basic tenet of the energy landscape model is that proteins fold through many heterogeneous pathways by way of amino acid-level dynamics biased toward selecting native-like interactions. The many pathways imagined in the model are not observed in the structure-formation stage of folding by experiments that would have found them, but they have now been detected and characterized for one protein in the initial prenucleation stage. Analysis presented here shows that these many microscopic trajectories are not distinct in any functionally significant way, and they have neither the structural information nor the biased energetics needed to select native vs. nonnative interactions during folding. The opposed defined-pathway model stems from experimental results that show that proteins are assemblies of small cooperative units called foldons and that a number of proteins fold in a reproducible pathway one foldon unit at a time. Thus, the same foldon interactions that encode the native structure of any given protein also naturally encode its particular foldon-based folding pathway, and they collectively sum to produce the energy bias toward native interactions that is necessary for efficient folding. Available information suggests that quantized native structure and stepwise folding coevolved in ancient repeat proteins and were retained as a functional pair due to their utility for solving the difficult protein folding problem.
Keywords: energy landscape theory; foldons; protein folding.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Clash between energy landscape theory and foldon-dependent protein folding.Proc Natl Acad Sci U S A. 2017 Aug 8;114(32):8442-8443. doi: 10.1073/pnas.1709133114. Epub 2017 Jul 26. Proc Natl Acad Sci U S A. 2017. PMID: 28747526 Free PMC article. No abstract available.
-
Theory, simulations, and experiments show that proteins fold by multiple pathways.Proc Natl Acad Sci U S A. 2017 Nov 14;114(46):E9759-E9760. doi: 10.1073/pnas.1716444114. Epub 2017 Oct 30. Proc Natl Acad Sci U S A. 2017. PMID: 29087352 Free PMC article. No abstract available.
-
Reply to Eaton and Wolynes: How do proteins fold?Proc Natl Acad Sci U S A. 2017 Nov 14;114(46):E9761-E9762. doi: 10.1073/pnas.1716929114. Epub 2017 Oct 30. Proc Natl Acad Sci U S A. 2017. PMID: 29087353 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
