

Mild metabolic perturbations alter succinylation of mitochondrial proteins
- PMID: 28631845
- PMCID: PMC5597472
- DOI: 10.1002/jnr.24103
Mild metabolic perturbations alter succinylation of mitochondrial proteins
Abstract
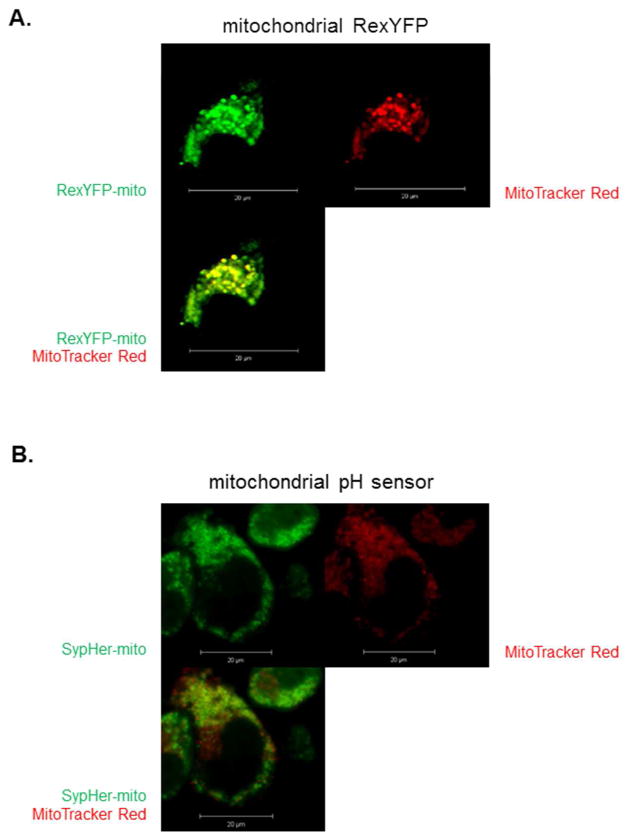
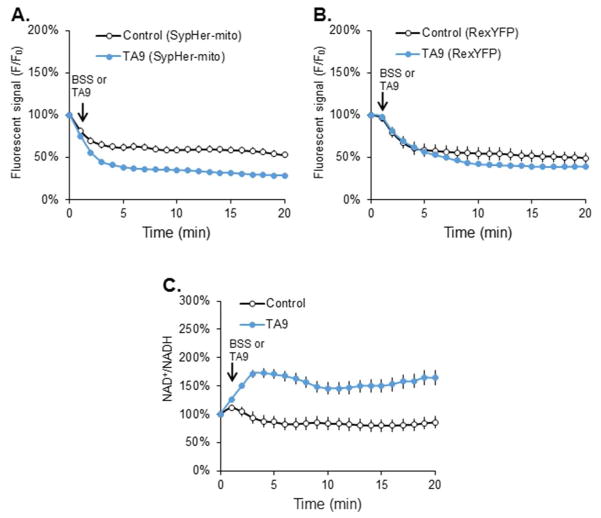
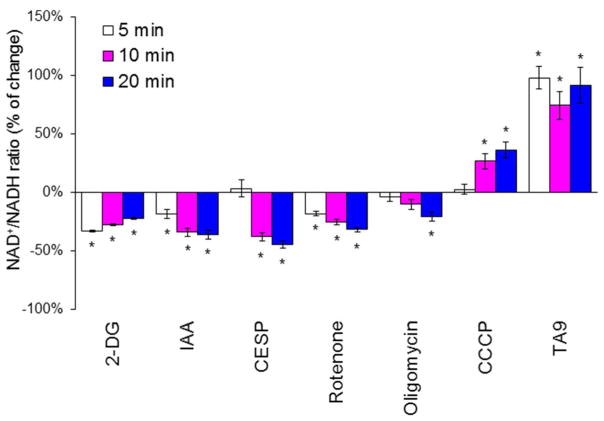
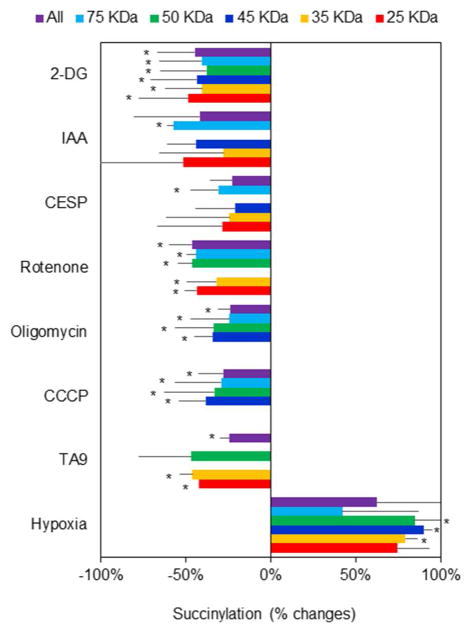
Succinylation of proteins is widespread, modifies both the charge and size of the molecules, and can alter their function. For example, liver mitochondrial proteins have 1,190 unique succinylation sites representing multiple metabolic pathways. Succinylation is sensitive to both increases and decreases of the NAD+ -dependent desuccinylase, SIRT5. Although the succinyl group for succinylation is derived from metabolism, the effects of systematic variation of metabolism on mitochondrial succinylation are not known. Changes in succinylation of mitochondrial proteins following variations in metabolism were compared against the mitochondrial redox state as estimated by the mitochondrial NAD+ /NADH ratio using fluorescent probes. The ratio was decreased by reduced glycolysis and/or glutathione depletion (iodoacetic acid; 2-deoxyglucose), depressed tricarboxylic acid cycle activity (carboxyethyl ester of succinyl phosphonate), and impairment of electron transport (antimycin) or ATP synthase (oligomycin), while uncouplers of oxidative phosphorylation (carbonyl cyanide m-chlorophenyl hydrazine or tyrphostin) increased the NAD+ /NADH ratio. All of the conditions decreased succinylation. In contrast, reducing the oxygen from 20% to 2.4% increased succinylation. The results demonstrate that succinylation varies with metabolic states, is not correlated to the mitochondrial NAD+ /NADH ratio, and may help coordinate the response to metabolic challenge.
Keywords: electron transport chain; glucose; hypoxia; succinylation; α-ketoglutarate dehydrogenase.
© 2017 Wiley Periodicals, Inc.
Figures
References
-
- Bunik VI, Denton TT, Xu H, Thompson CM, Cooper AJL, Gibson GE. Phosphonate Analogues of α-Ketoglutarate Inhibit the Activity of the α-Ketoglutarate Dehydrogenase Complex Isolated from Brain and in Cultured Cells. Biochemistry. 2005;44(31):10552–10561. - PubMed
-
- Choudhary C, Weinert BT, Nishida Y, Verdin E, Mann M. The growing landscape of lysine acetylation links metabolism and cell signalling. Nat Rev Mol Cell Biol. 2014;15(8):536–550. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
