

Structural analyses of human thymidylate synthase reveal a site that may control conformational switching between active and inactive states
- PMID: 28634233
- PMCID: PMC5555203
- DOI: 10.1074/jbc.M117.787267
Structural analyses of human thymidylate synthase reveal a site that may control conformational switching between active and inactive states
Abstract
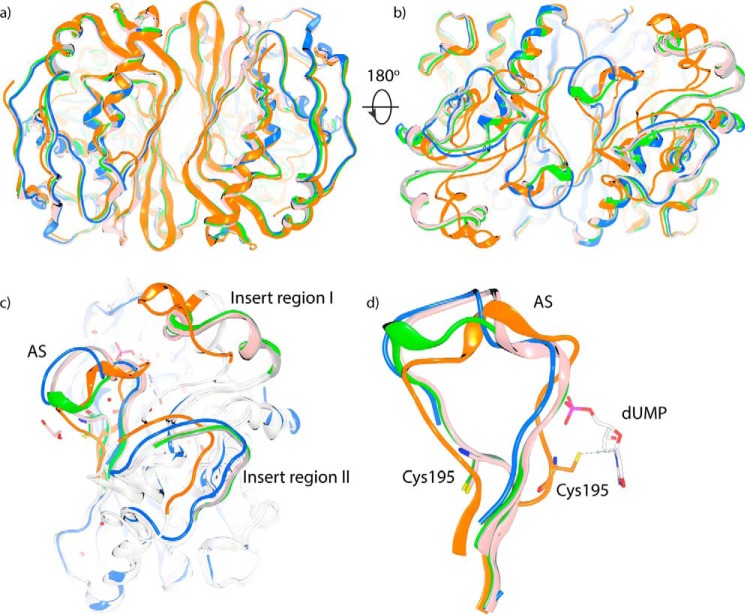
Thymidylate synthase (TS) is the sole enzyme responsible for de novo biosynthesis of thymidylate (TMP) and is essential for cell proliferation and survival. Inhibition of human TS (hTS) has been extensively investigated for cancer chemotherapy, but several aspects of its activity and regulation are still uncertain. In this study, we performed comprehensive structural and biophysical studies of hTS using crystallography and thermal shift assay and provided the first detailed structural information on the conformational changes induced by ligand binding to the hTS active site. We found that upon binding of the antifolate agents raltitrexed and nolatrexed, the two insert regions in hTS, the functions of which are unclear, undergo positional shifts toward the catalytic center. We investigated the inactive conformation of hTS and found that the two insert regions are also involved in the conformational transition between the active and inactive state of hTS. Moreover, we identified a ligand-binding site in the dimer interface, suggesting that the cavity in the dimer interface could serve as an allosteric site of hTS to regulate the conformational switching between the active and inactive states. On the basis of these findings, we propose a regulatory mechanism of hTS activity that involves allosteric regulation of interactions of hTS with its own mRNA depending on cellular demands for TMP.
Keywords: DNA synthesis; antifolate; conformational change; crystallography; drug resistance; nucleotide; thymidylate synthase.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures




References
-
- Hori T., Ayusawa D., Shimizu K., Koyama H., and Seno T. (1984) Chromosome breakage induced by thymidylate stress in thymidylate synthase-negative mutants of mouse FM3A cells. Cancer Res. 44, 703–709 - PubMed
-
- Seno T., Ayusawa D., Shimizu K., Koyama H., Takeishi K., and Hori T. (1985) Thymineless death and genetic events in mammalian cells. Basic Life Sci. 31, 241–263 - PubMed
-
- Wilson P. M., Danenberg P. V., Johnston P. G., Lenz H. J., and Ladner R. D. (2014) Standing the test of time: targeting thymidylate biosynthesis in cancer therapy. Nat. Rev. Clin. Oncol. 11, 282–298 - PubMed
-
- Ackland S. P., Clarke S. J., Beale P., and Peters G. J. (2006) Thymidylate synthase inhibitors. Update Cancer Therap. 1, 403–427 - PubMed
-
- Assaraf Y. (2007) Molecular basis of antifolate resistance. Cancer Metastasis Rev. 26, 153–181 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
