

Characterization of the Proinflammatory Profile of Synovial Fluid-Derived Exosomes of Patients with Osteoarthritis
- PMID: 28634420
- PMCID: PMC5467328
- DOI: 10.1155/2017/4814987
Characterization of the Proinflammatory Profile of Synovial Fluid-Derived Exosomes of Patients with Osteoarthritis
Abstract
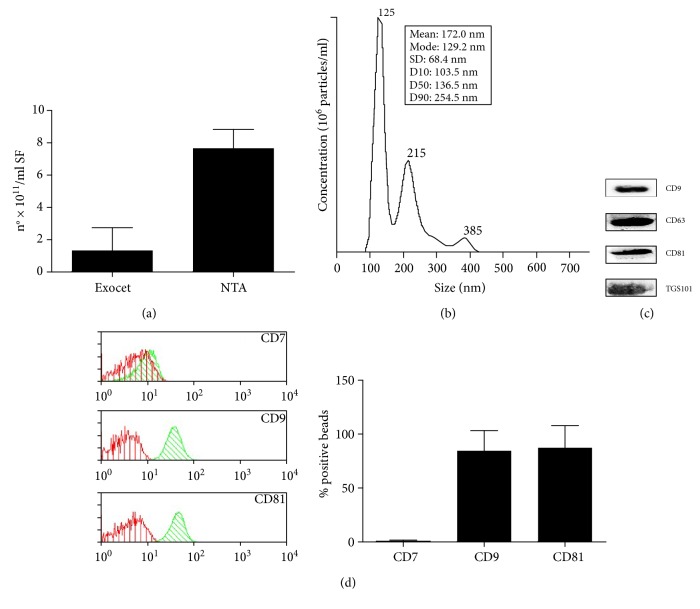
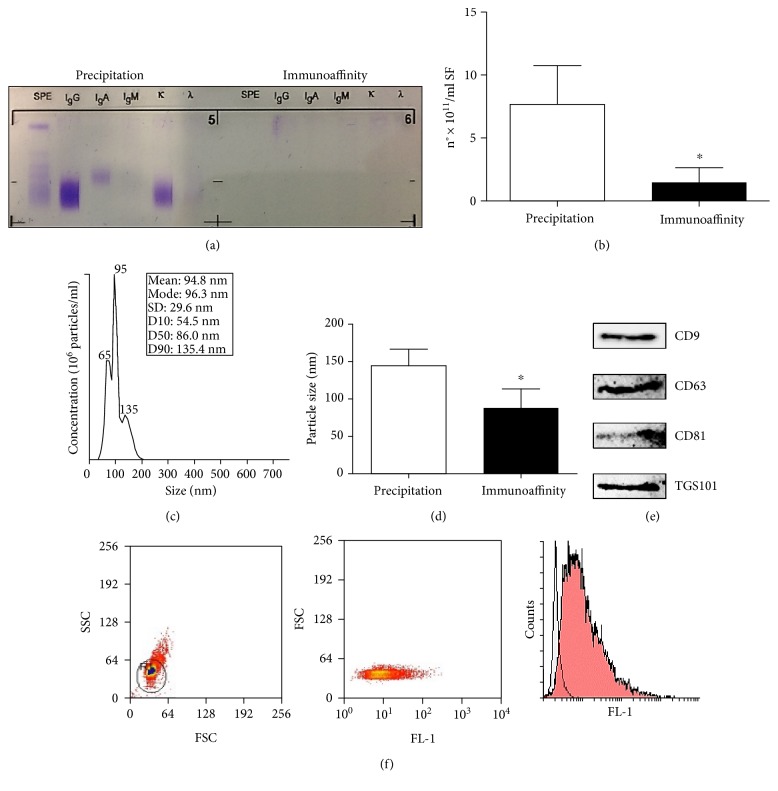
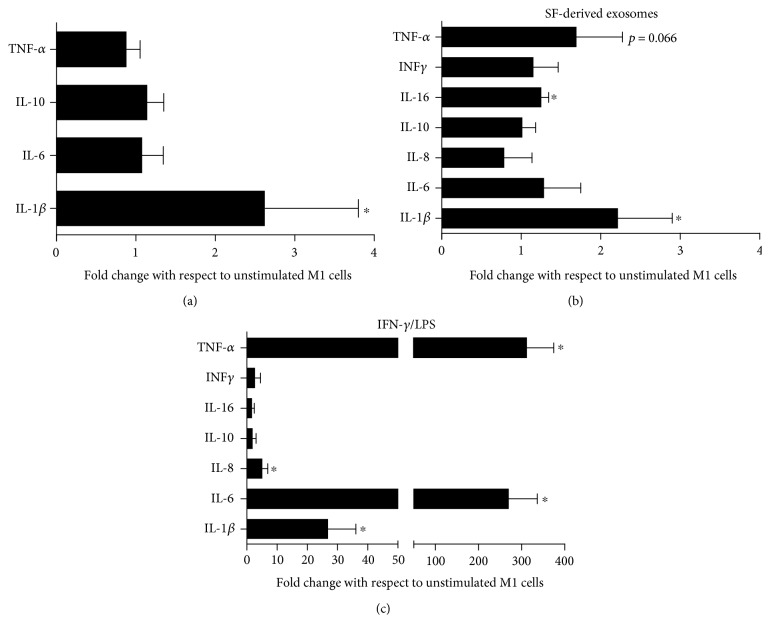
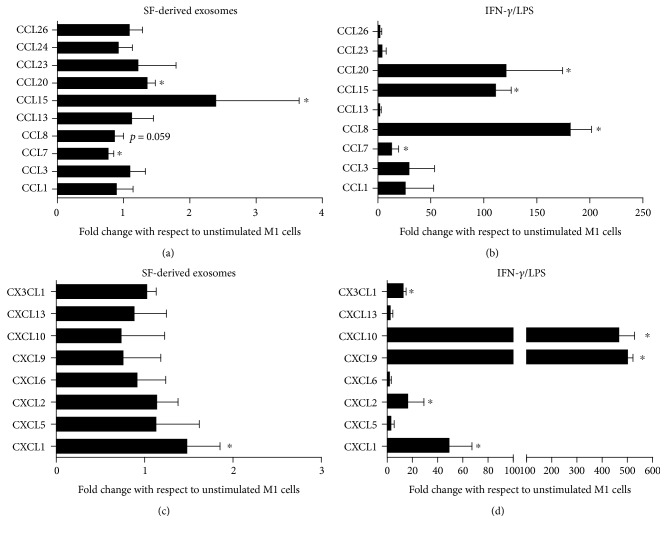
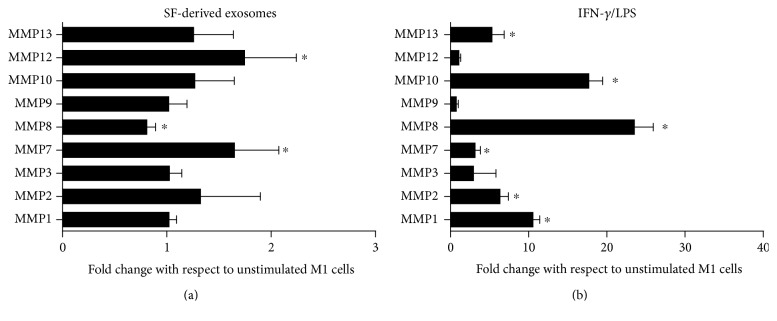
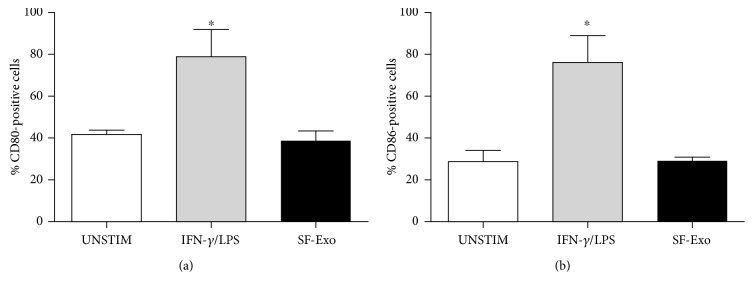
The purpose of this study is to characterize synovial fluid- (SF-) derived exosomes of patients with gonarthrosis comparing two methods of isolation and to investigate their immune regulatory properties. Extracellular vesicles (EVs) have been isolated from inflamed SF by polymer precipitation method and quantified by Exocet kit and by nanoparticle tracking analysis. Vesicles expressed all the specific exosomal markers by immunoblot and FACS. After isolation with Exoquick, a relevant contamination by immune complexes was detected, which required further magnetic bead-based purification to remove. SF-derived exosomes significantly stimulated the release of several inflammatory cytokines and chemokines and metalloproteinases by M1 macrophages but did not influence the expression of CD80 and CD86 costimulatory molecules. In conclusion, we characterized purified exosomes isolated from inflamed SF and demonstrate that purified exosomes are functionally active in their ability to stimulate the release of proinflammatory factors from M1 macrophages. Our data indicate that SF-derived exosomes from gonarthrosis patients play a role in disease progression.
Figures
References
-
- Bondeson J., Blom A. B., Wainwright S., Hughes C., Caterson B., van den Berg W. B. The role of synovial macrophages and macrophage-produced mediators in driving inflammatory and destructive responses in osteoarthritis. Arthritis and Rheumatism. 2010;62(3):647–657. doi: 10.1002/art.27290. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
