

Critical biological parameters modulate affinity as a determinant of function in T-cell receptor gene-modified T-cells
- PMID: 28634816
- PMCID: PMC5647210
- DOI: 10.1007/s00262-017-2032-9
Critical biological parameters modulate affinity as a determinant of function in T-cell receptor gene-modified T-cells
Abstract
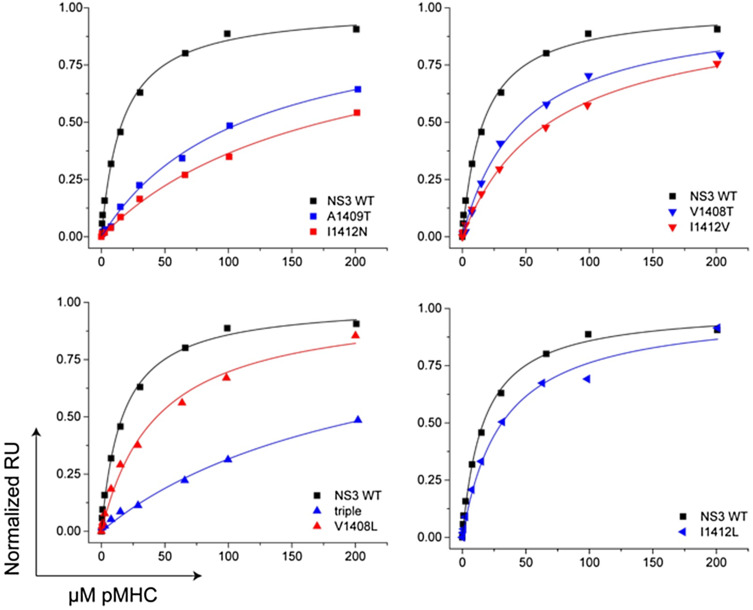
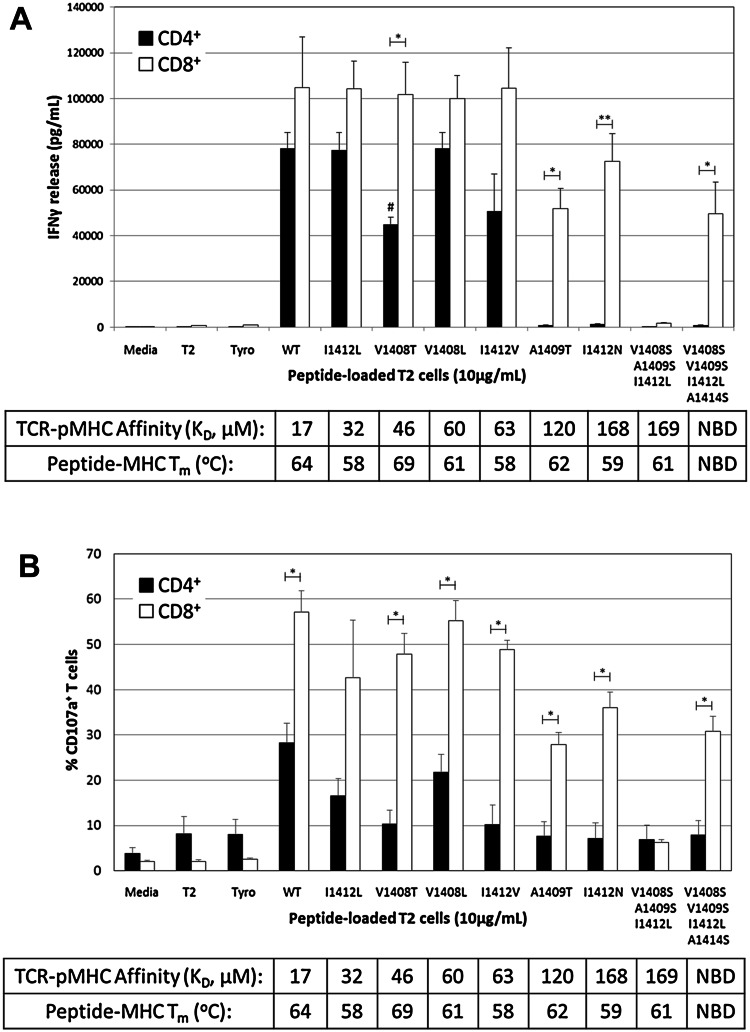
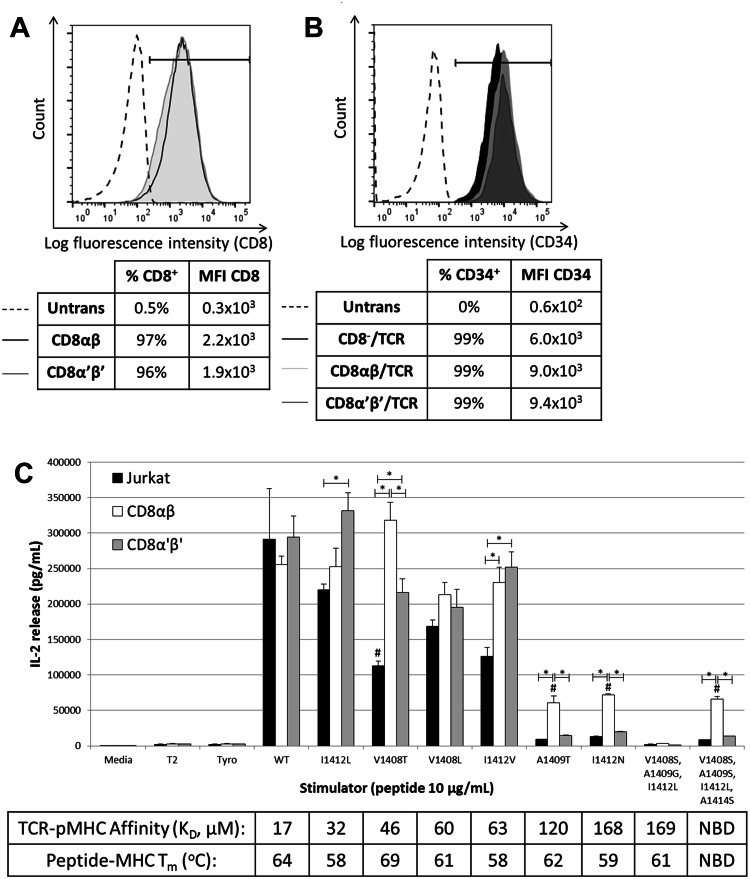
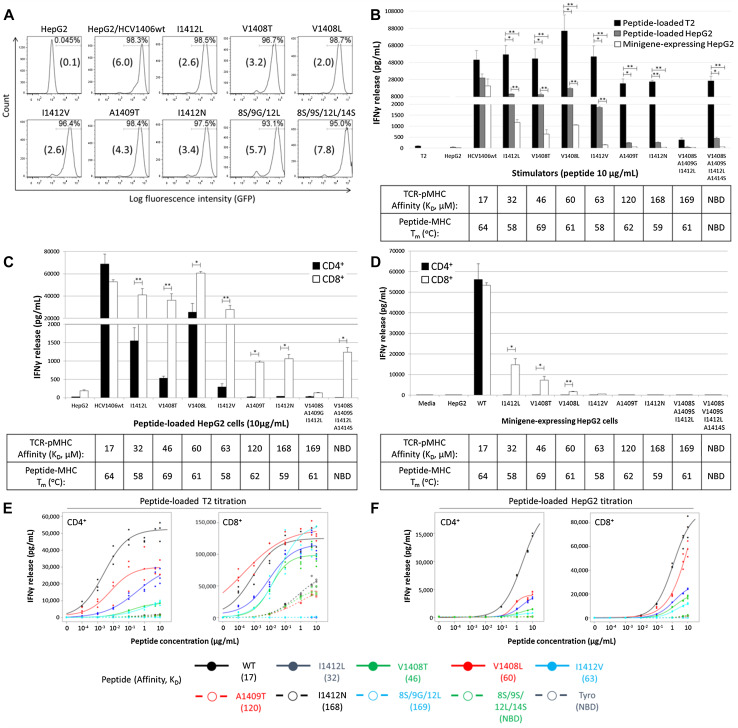
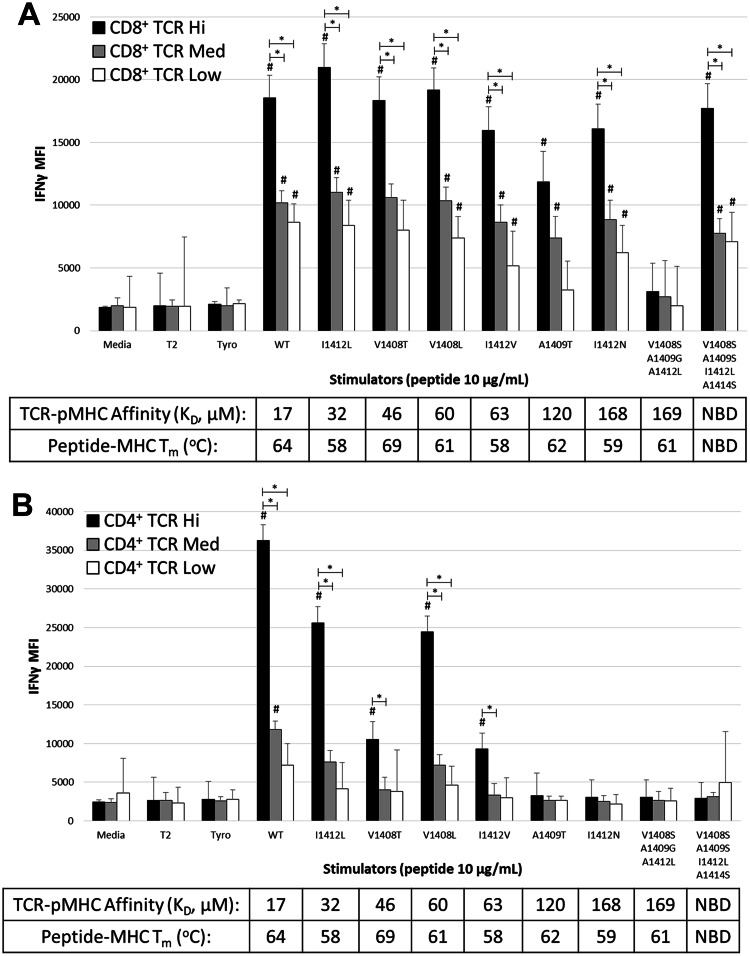
T-cell receptor (TCR)-pMHC affinity has been generally accepted to be the most important factor dictating antigen recognition in gene-modified T-cells. As such, there is great interest in optimizing TCR-based immunotherapies by enhancing TCR affinity to augment the therapeutic benefit of TCR gene-modified T-cells in cancer patients. However, recent clinical trials using affinity-enhanced TCRs in adoptive cell transfer (ACT) have observed unintended and serious adverse events, including death, attributed to unpredicted off-tumor or off-target cross-reactivity. It is critical to re-evaluate the importance of other biophysical, structural, or cellular factors that drive the reactivity of TCR gene-modified T-cells. Using a model for altered antigen recognition, we determined how TCR-pMHC affinity influenced the reactivity of hepatitis C virus (HCV) TCR gene-modified T-cells against a panel of naturally occurring HCV peptides and HCV-expressing tumor targets. The impact of other factors, such as TCR-pMHC stabilization and signaling contributions by the CD8 co-receptor, as well as antigen and TCR density were also evaluated. We found that changes in TCR-pMHC affinity did not always predict or dictate IFNγ release or degranulation by TCR gene-modified T-cells, suggesting that less emphasis might need to be placed on TCR-pMHC affinity as a means of predicting or augmenting the therapeutic potential of TCR gene-modified T-cells used in ACT. A more complete understanding of antigen recognition by gene-modified T-cells and a more rational approach to improve the design and implementation of novel TCR-based immunotherapies is necessary to enhance efficacy and maximize safety in patients.
Keywords: Adoptive cell therapy; Affinity; Altered peptide ligands; Gene-modified T-cells; T-cell; T-cell receptor (TCR).
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Clay TM, Custer MC, Sachs J, Hwu P, Rosenberg SA, Nishimura MI. Efficient transfer of a tumor antigen-reactive TCR to human peripheral blood lymphocytes confers anti-tumor reactivity. J Immunol. 1999;163(1):507–513. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
