

Pyridine Dinucleotides from Molecules to Man
- PMID: 28635300
- PMCID: PMC5737636
- DOI: 10.1089/ars.2017.7120
Pyridine Dinucleotides from Molecules to Man
Abstract
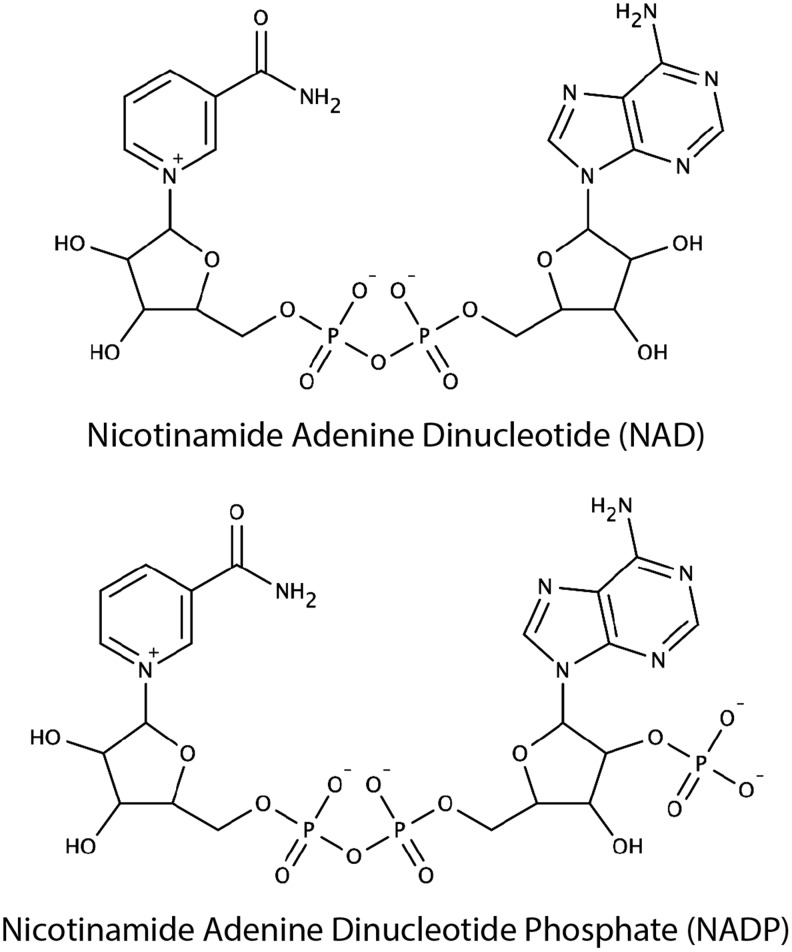
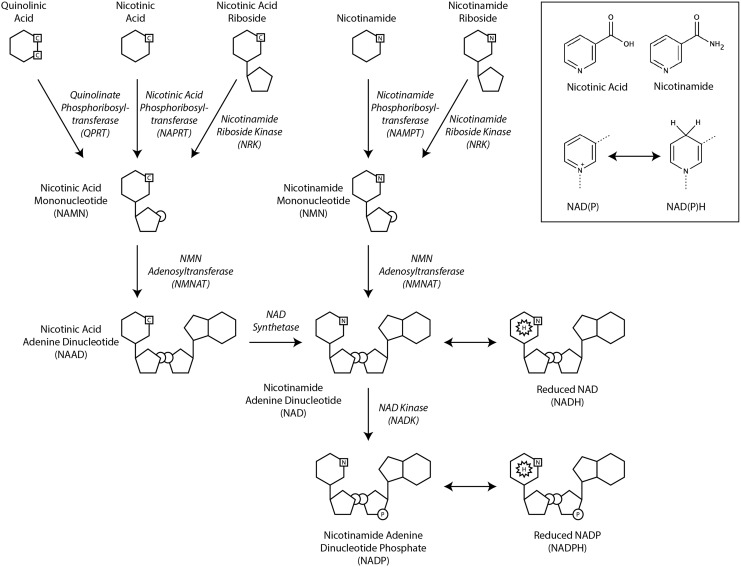
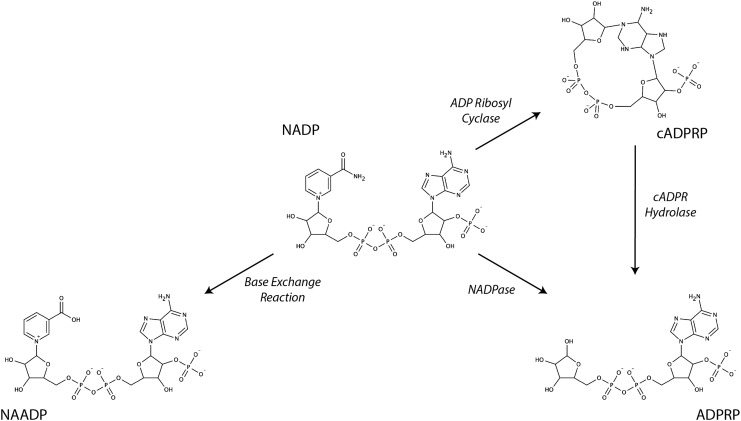
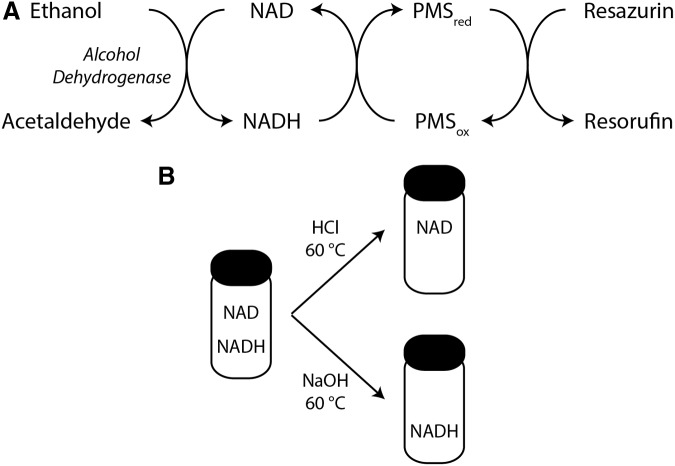
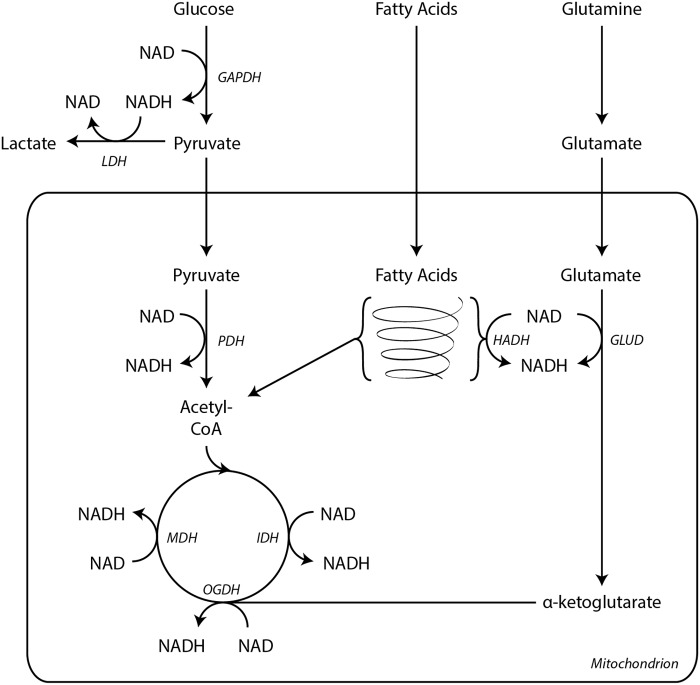
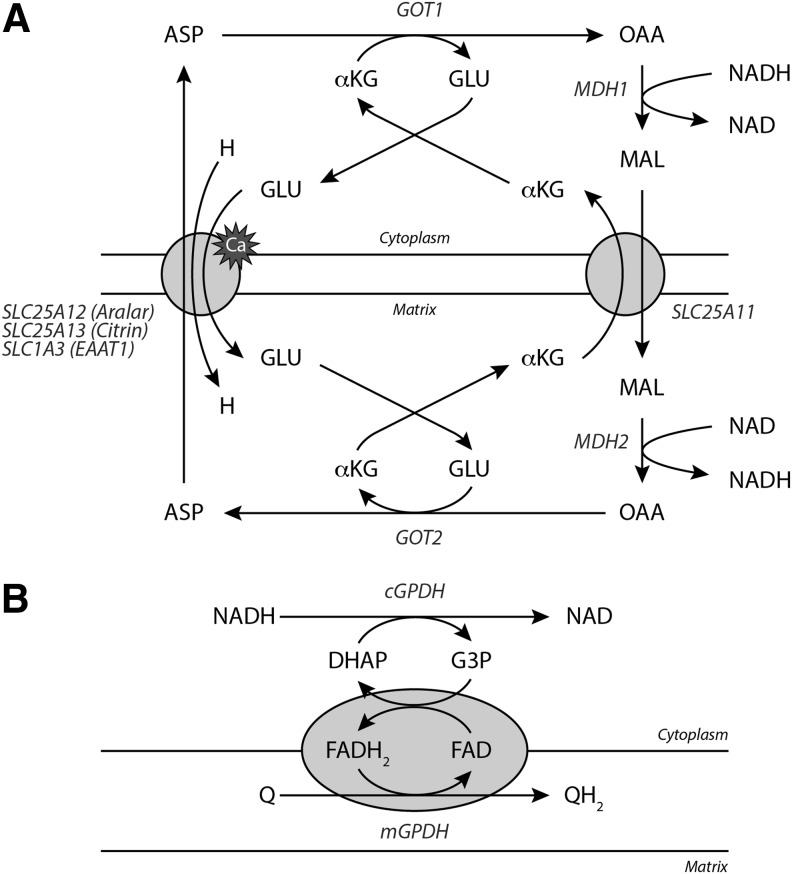
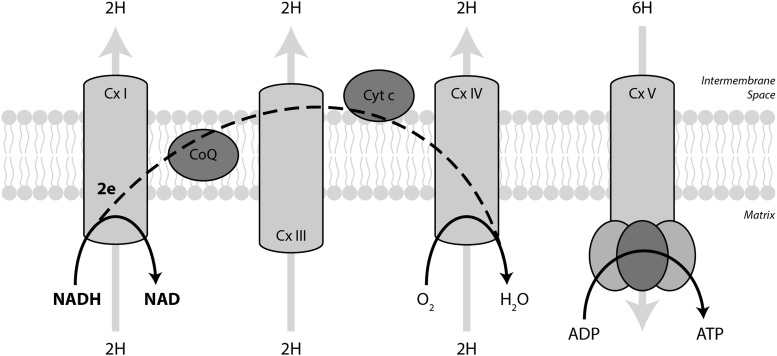
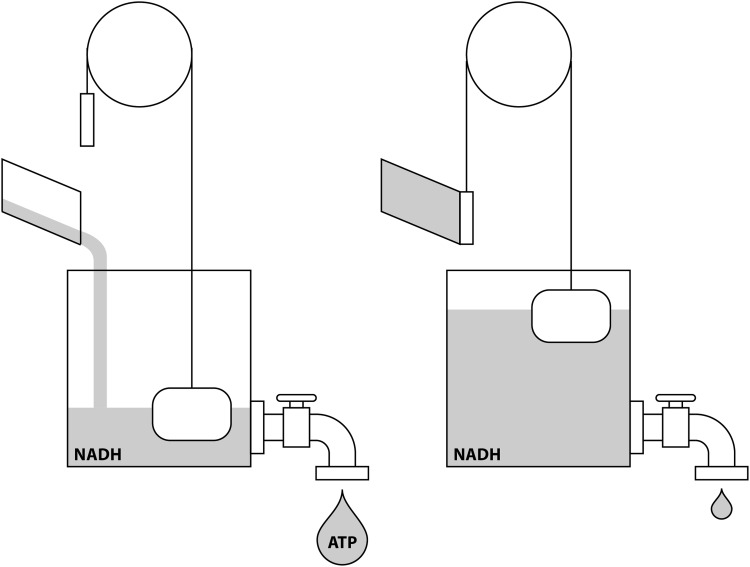
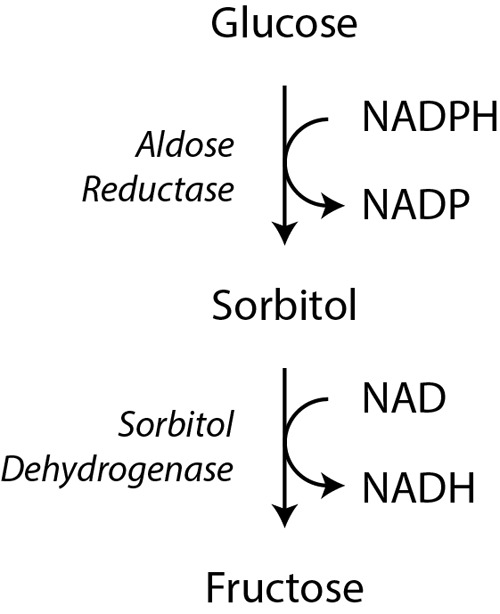
Significance: Pyridine dinucleotides, nicotinamide adenine dinucleotide (NAD) and nicotinamide adenine dinucleotide phosphate (NADP), were discovered more than 100 years ago as necessary cofactors for fermentation in yeast extracts. Since that time, these molecules have been recognized as fundamental players in a variety of cellular processes, including energy metabolism, redox homeostasis, cellular signaling, and gene transcription, among many others. Given their critical role as mediators of cellular responses to metabolic perturbations, it is unsurprising that dysregulation of NAD and NADP metabolism has been associated with the pathobiology of many chronic human diseases. Recent Advances: A biochemistry renaissance in biomedical research, with its increasing focus on the metabolic pathobiology of human disease, has reignited interest in pyridine dinucleotides, which has led to new insights into the cell biology of NAD(P) metabolism, including its cellular pharmacokinetics, biosynthesis, subcellular localization, and regulation. This review highlights these advances to illustrate the importance of NAD(P) metabolism in the molecular pathogenesis of disease.
Critical issues: Perturbations of NAD(H) and NADP(H) are a prominent feature of human disease; however, fundamental questions regarding the regulation of the absolute levels of these cofactors and the key determinants of their redox ratios remain. Moreover, an integrated topological model of NAD(P) biology that combines the metabolic and other roles remains elusive.
Future directions: As the complex regulatory network of NAD(P) metabolism becomes illuminated, sophisticated new approaches to manipulating these pathways in specific organs, cells, or organelles will be developed to target the underlying pathogenic mechanisms of disease, opening doors for the next generation of redox-based, metabolism-targeted therapies. Antioxid. Redox Signal. 28, 180-212.
Keywords: NAD; NADP; PARP; SIRT; metabolism.
Figures

References
-
- Institute of Medicine. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline. Washington, DC: The National Academies Press; 1998, p. 592 - PubMed
-
- Alano CC, Tran A, Tao R, Ying W, Karliner JS, and Swanson RA. Differences among cell types in NAD(+) compartmentalization: a comparison of neurons, astrocytes, and cardiac myocytes. J Neurosci Res 85: 3378–3385, 2007 - PubMed
-
- Avalos JL, Bever KM, and Wolberger C. Mechanism of sirtuin inhibition by nicotinamide: altering the NAD(+) cosubstrate specificity of a Sir2 enzyme. Mol Cell 17: 855–868, 2005 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
