

Family living sets the stage for cooperative breeding and ecological resilience in birds
- PMID: 28636615
- PMCID: PMC5479502
- DOI: 10.1371/journal.pbio.2000483
Family living sets the stage for cooperative breeding and ecological resilience in birds
Abstract
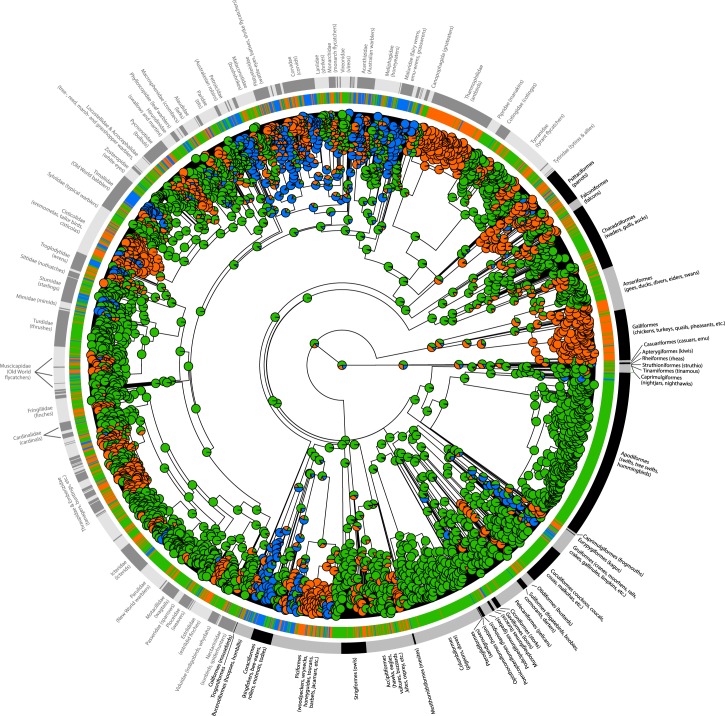
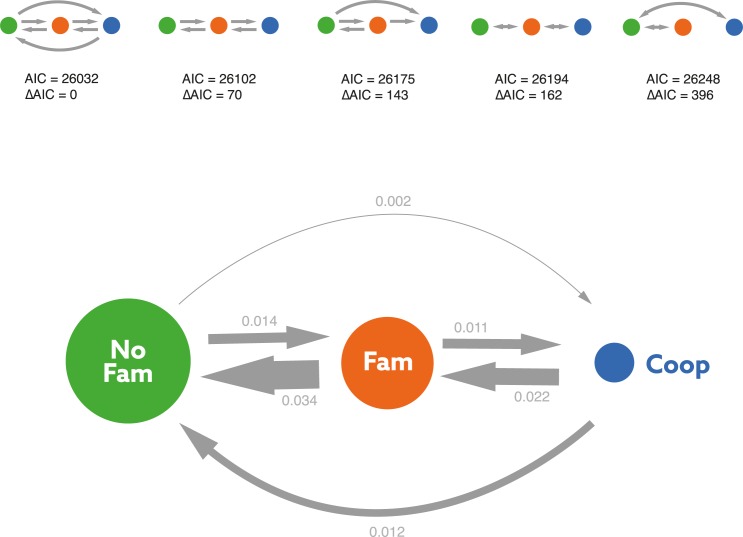
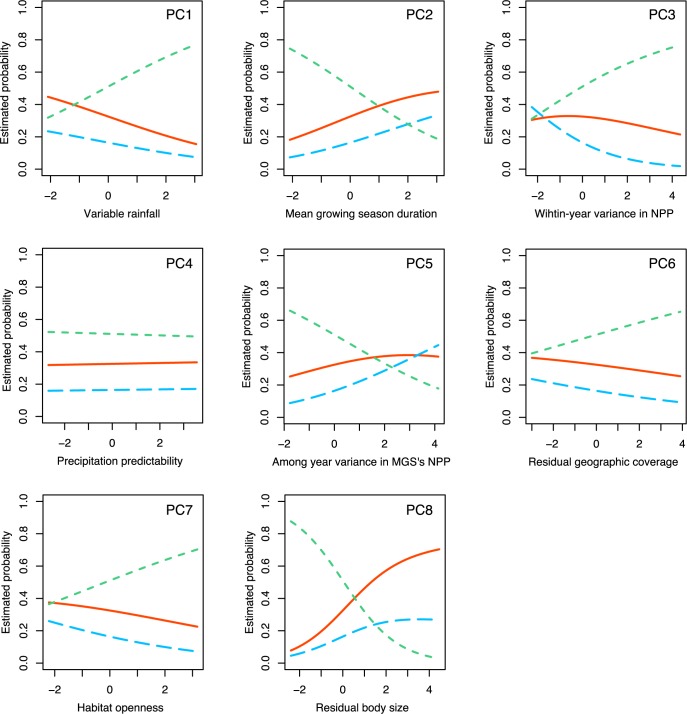
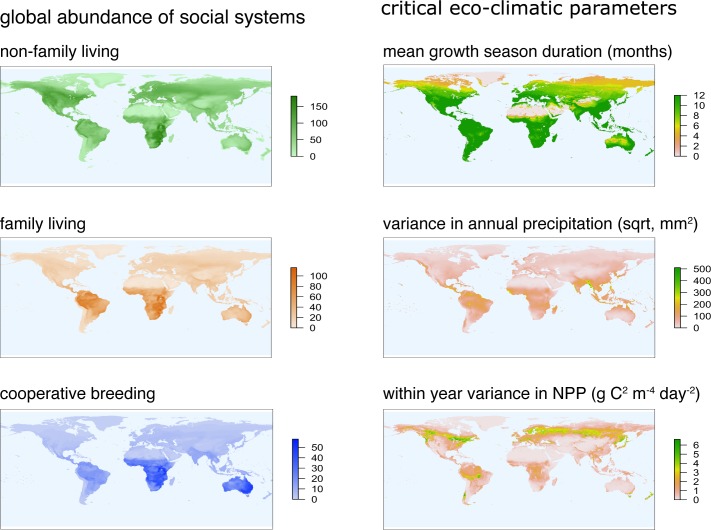
Cooperative breeding is an extreme form of cooperation that evolved in a range of lineages, including arthropods, fish, birds, and mammals. Although cooperative breeding in birds is widespread and well-studied, the conditions that favored its evolution are still unclear. Based on phylogenetic comparative analyses on 3,005 bird species, we demonstrate here that family living acted as an essential stepping stone in the evolution of cooperative breeding in the vast majority of species. First, families formed by prolonging parent-offspring associations beyond nutritional independency, and second, retained offspring began helping at the nest. These findings suggest that assessment of the conditions that favor the evolution of cooperative breeding can be confounded if this process is not considered to include 2 steps. Specifically, phylogenetic linear mixed models show that the formation of families was associated with more productive and seasonal environments, where prolonged parent-offspring associations are likely to be less costly. However, our data show that the subsequent evolution of cooperative breeding was instead linked to environments with variable productivity, where helpers at the nest can buffer reproductive failure in harsh years. The proposed 2-step framework helps resolve current disagreements about the role of environmental forces in the evolution of cooperative breeding and better explains the geographic distribution of this trait. Many geographic hotspots of cooperative breeding have experienced a historical decline in productivity, suggesting that a higher proportion of family-living species could have been able to avoid extinction under harshening conditions through the evolution of cooperative breeding. These findings underscore the importance of considering the potentially different factors that drive different steps in the evolution of complex adaptations.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Comment in
-
Birds, blooms, and evolving diversity.PLoS Biol. 2018 Oct 4;16(10):e3000020. doi: 10.1371/journal.pbio.3000020. eCollection 2018 Oct. PLoS Biol. 2018. PMID: 30286070 Free PMC article.
References
-
- Griesser M, Suzuki TN. Occasional cooperative breeding in birds and the robustness of comparative analyses concerning the evolution of cooperative breeding. Zoological Letters. 2016:2:7 doi: 10.1186/s40851-016-0041-8 - DOI - PMC - PubMed
-
- Koenig WD, Dickinson J. Cooperative Breeding in Vertebrates: Studies of Ecology, Evolution, and Behavior Cambridge: Cambridge University Press; 2016.
-
- Emlen ST. The Evolution of Helping .1. an Ecological Constraints Model. American Naturalist. 1982;119(1):29–39. .
-
- Darwin C. On the origin of species. London: John Murray; 1859.
-
- Hamilton WD. Genetical Evolution of Social Behaviour I. Journal of Theoretical Biology. 1964;7(1):1–16. . - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
