

The Super-Enhancer-Derived alncRNA-EC7/Bloodlinc Potentiates Red Blood Cell Development in trans
- PMID: 28636939
- PMCID: PMC6013260
- DOI: 10.1016/j.celrep.2017.05.082
The Super-Enhancer-Derived alncRNA-EC7/Bloodlinc Potentiates Red Blood Cell Development in trans
Abstract
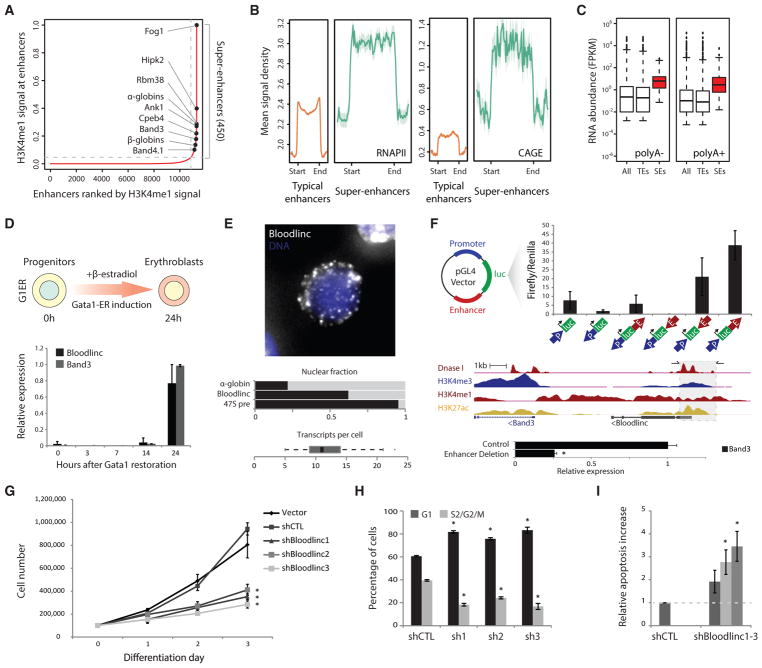
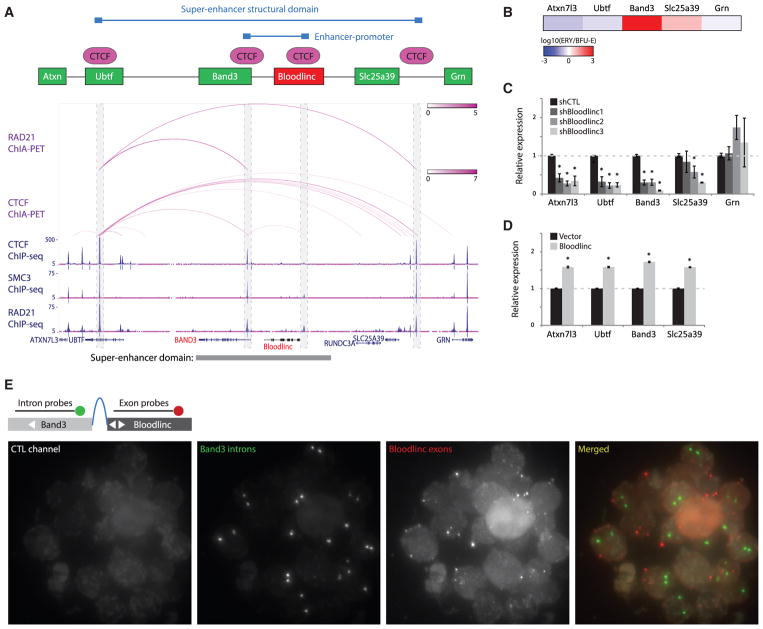
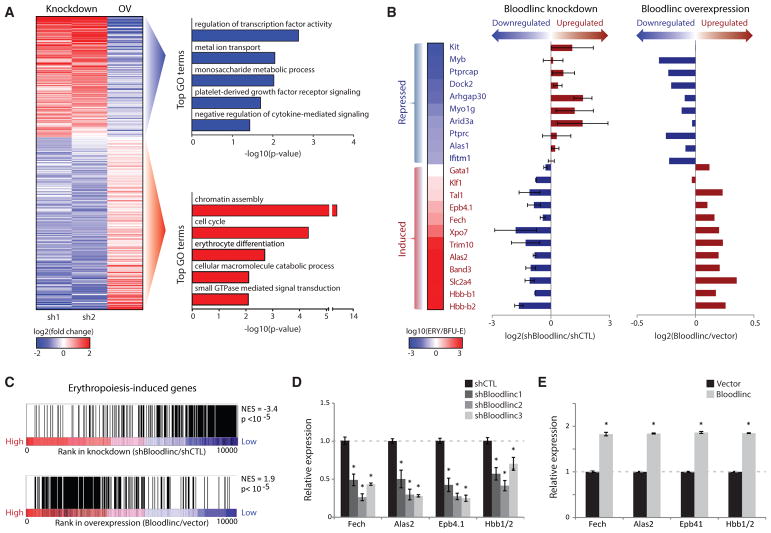
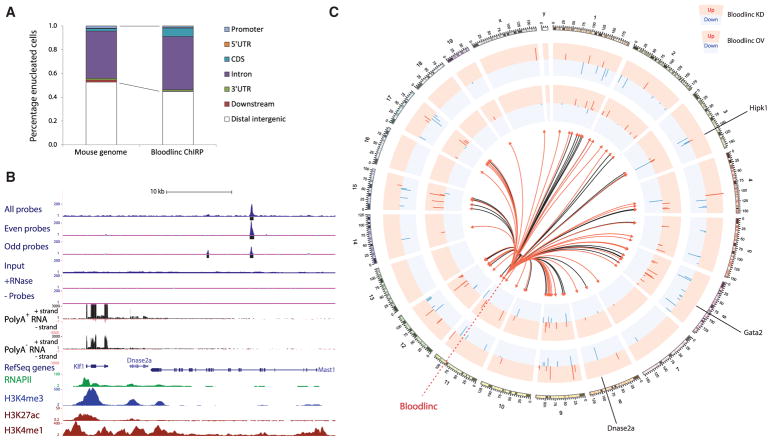
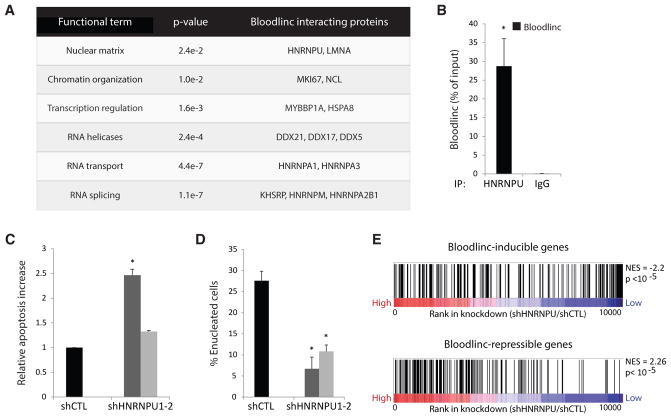
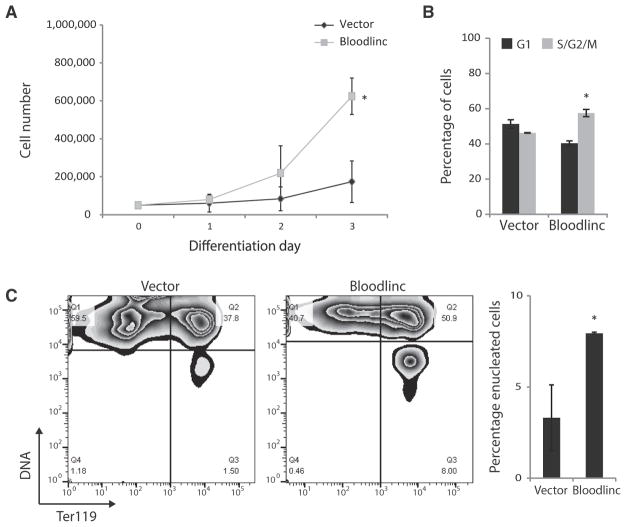
Enhancer-derived RNAs are thought to act locally by contributing to their parent enhancer function. Whether large domains of clustered enhancers (super-enhancers) also produce cis-acting RNAs, however, remains unclear. Unlike typical enhancers, super-enhancers form large spans of robustly transcribed chromatin, amassing capped and polyadenylated RNAs that are sufficiently abundant to sustain trans functions. Here, we show that one such RNA, alncRNA-EC7/Bloodlinc, is transcribed from a super-enhancer of the erythroid membrane transporter SLC4A1/BAND3 but diffuses beyond this site. Bloodlinc localizes to trans-chromosomal loci encoding critical regulators and effectors of terminal erythropoiesis and directly binds chromatin-organizing and transcription factors, including the chromatin attachment factor HNRNPU. Inhibiting Bloodlinc or Hnrnpu compromises the terminal erythropoiesis gene program, blocking red cell production, whereas expressing Bloodlinc ectopically stimulates this program and can promote erythroblast proliferation and enucleation in the absence of differentiation stimuli. Thus, Bloodlinc is a trans-acting super-enhancer RNA that potentiates red blood cell development.
Keywords: enhancer RNA; erythropoiesis; long non-coding RNA; red blood cell; super-enhancer.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
On the Origin of lncRNAs: Missing Link Found.Trends Genet. 2017 Oct;33(10):660-662. doi: 10.1016/j.tig.2017.07.005. Epub 2017 Aug 1. Trends Genet. 2017. PMID: 28778681 Free PMC article.
References
-
- Burke B, Stewart CL. The nuclear lamins: flexibility in function. Nat Rev Mol Cell Biol. 2013;14:13–24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
