

Identification of an Intermediate in Hepatitis B Virus Covalently Closed Circular (CCC) DNA Formation and Sensitive and Selective CCC DNA Detection
- PMID: 28637752
- PMCID: PMC5553186
- DOI: 10.1128/JVI.00539-17
Identification of an Intermediate in Hepatitis B Virus Covalently Closed Circular (CCC) DNA Formation and Sensitive and Selective CCC DNA Detection
Abstract
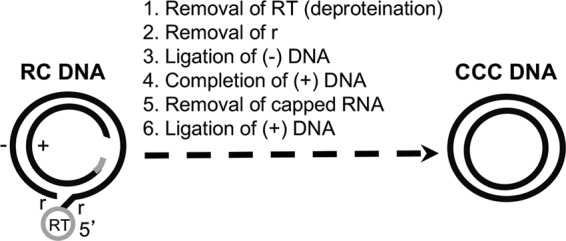
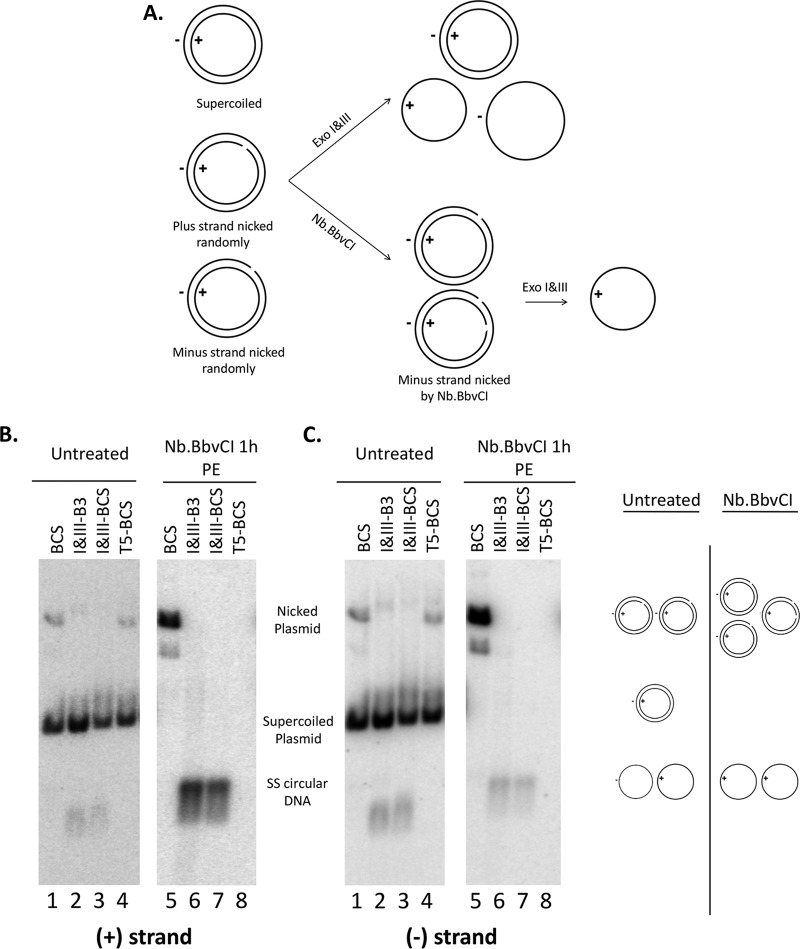
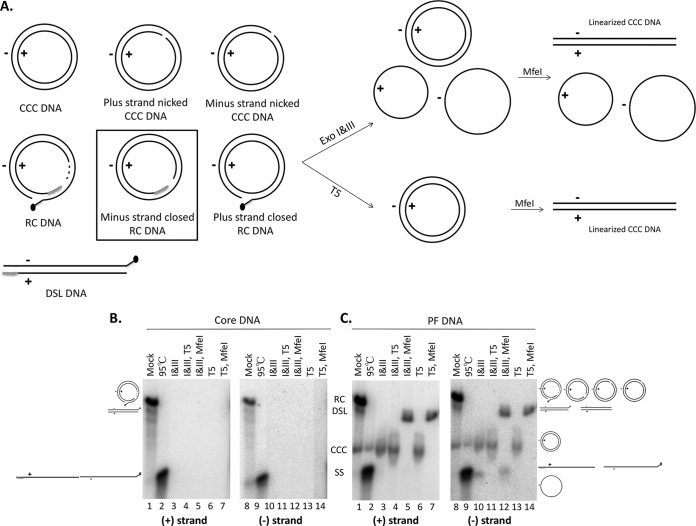
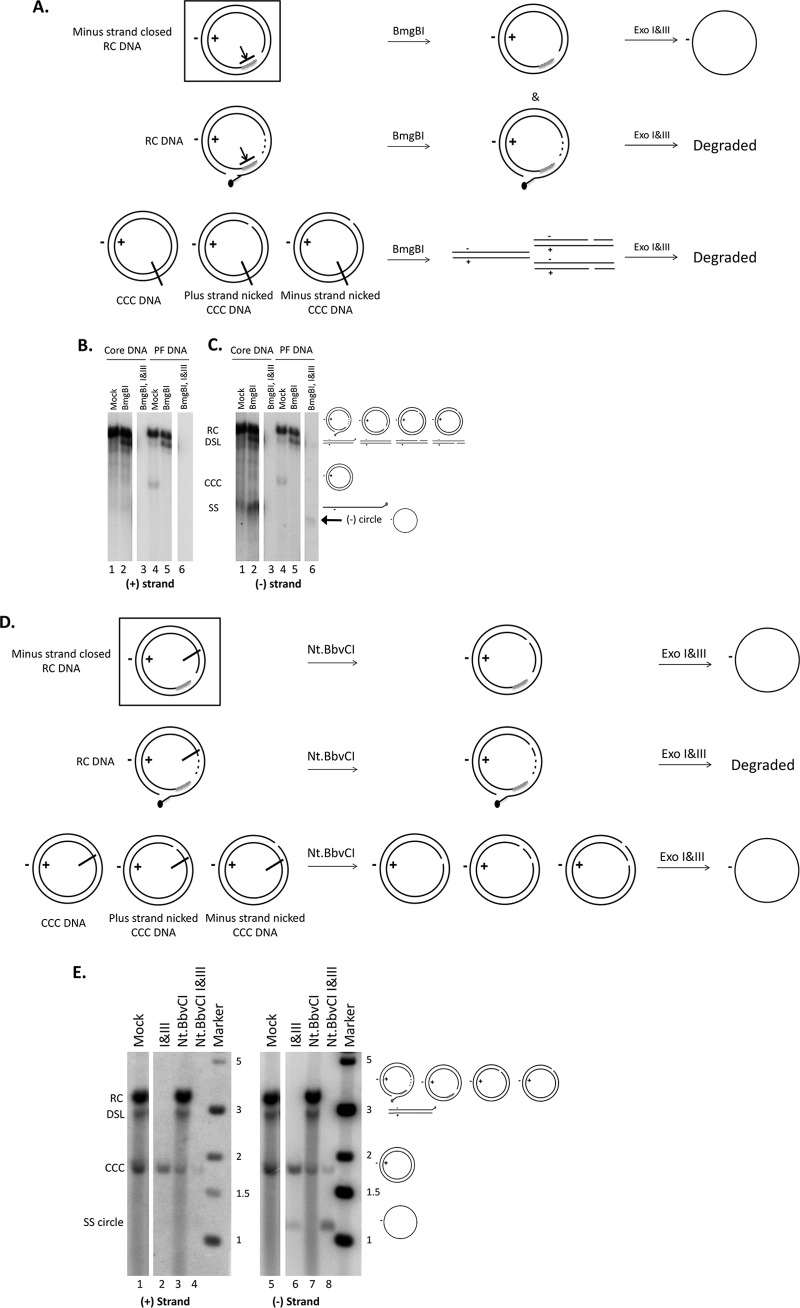
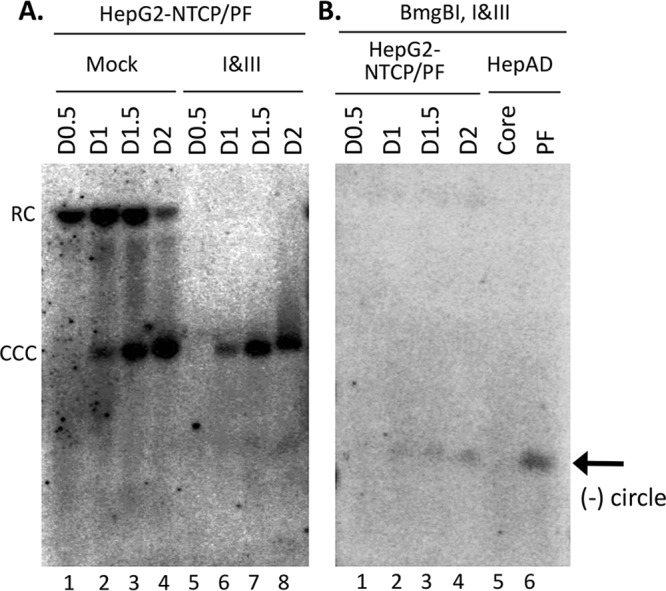
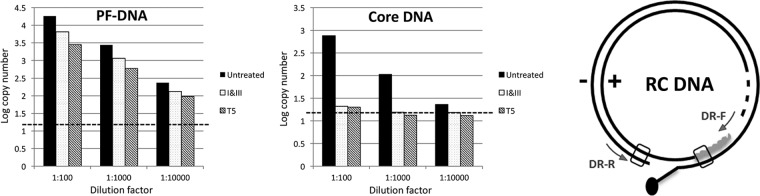
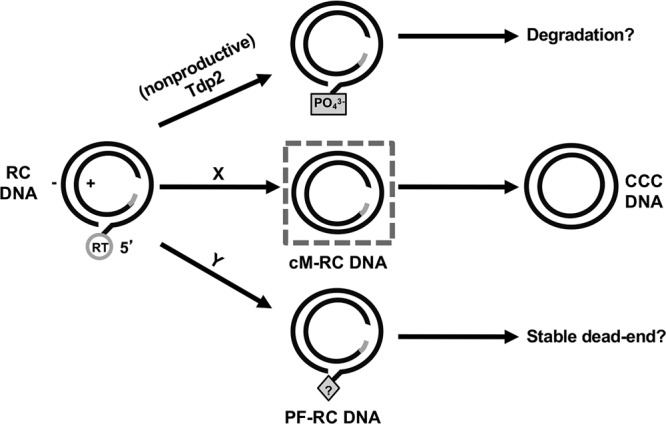
Hepatitis B virus (HBV) covalently closed circular (CCC) DNA functions as the only viral template capable of coding for all the viral RNA species and is thus essential to initiate and sustain viral replication. CCC DNA is converted, in a multistep and ill-understood process, from a relaxed circular (RC) DNA, in which neither of the two DNA strands is covalently closed. To detect putative intermediates during RC DNA to CCC DNA conversion, two 3' exonucleases, exonuclease I (Exo I) and Exo III, were used in combination to degrade all DNA strands with a free 3' end, which would nevertheless preserve closed circular DNA in either single-stranded (SS) or double-stranded (DS) form. Indeed, an RC DNA species with a covalently closed minus strand but an open plus strand (closed minus-strand RC DNA [cM-RC DNA]) was detected by this approach. Further analyses indicated that at least some of the plus strands in such a putative intermediate likely still retained the RNA primer that is attached to the 5' end of the plus strand in RC DNA, suggesting that minus-strand closing can occur before plus-strand processing. Furthermore, the same nuclease treatment proved to be useful for sensitive and specific detection of CCC DNA by removing all DNA species other than closed circular DNA. Application of these and similar approaches may allow the identification of additional intermediates during CCC DNA formation and facilitate specific and sensitive detection of CCC DNA, which should help elucidate the pathways of CCC DNA formation and the factors involved.IMPORTANCE The hepatitis B virus (HBV) covalently closed circular (CCC) DNA, by serving as the viral transcriptional template, is the molecular basis of viral persistence. CCC DNA is converted, in a multistep and ill-understood process, from relaxed circular (RC) DNA. Little is currently understood about the pathways or factors involved in CCC DNA formation. We have now detected a likely intermediate during the conversion of RC DNA to CCC DNA, thus providing important clues to the pathways of CCC DNA formation. Furthermore, the same experimental approach that led to the detection of the intermediate could also facilitate specific and sensitive detection of CCC DNA, which has remained challenging. This and similar approaches will help identify additional intermediates during CCC DNA formation and elucidate the pathways and factors involved.
Keywords: covalently closed circular DNA; exonuclease; hepatitis B virus; intermediate; relaxed circular DNA.
Copyright © 2017 American Society for Microbiology.
Figures

References
-
- Lavanchy D, Kane M. 2016. Global epidemiology of hepatitis B virus infection, p 187–203. In Liaw Y-F, Zoulim F (ed), Hepatitis B virus in human diseases. Humana Press, New York, NY.
-
- Hu J. 2016. Hepatitis B virus virology and replication, p 1–34. In Liaw Y-F, Zoulim F (ed), Hepatitis B virus in human diseases. Humana Press, New York, NY.
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
