

Review
doi: 10.1038/nrn.2017.61.
Epub 2017 Jun 22.
Neuroimmunology and neuroepigenetics in the establishment of sex differences in the brain
Affiliations
- PMID: 28638119
- PMCID: PMC5771241
- DOI: 10.1038/nrn.2017.61
Item in Clipboard
Review
Neuroimmunology and neuroepigenetics in the establishment of sex differences in the brain
Nat Rev Neurosci.
2017 Aug.
Abstract
The study of sex differences in the brain is a topic of neuroscientific study that has broad reaching implications for culture, society and biomedical science. Recent research in rodent models has led to dramatic shifts in our views of the mechanisms underlying the sexual differentiation of the brain. These include the surprising discoveries of a role for immune cells and inflammatory mediators in brain masculinization and a role for epigenetic suppression in brain feminization. How and to what degree these findings will translate to human brain development will be questions of central importance in future research in this field.
Conflict of interest statement
The authors declare no competing interests.
Figures
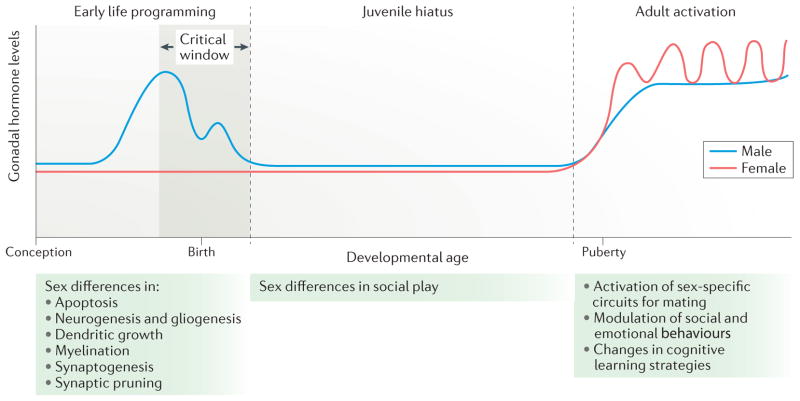
The role of hormones in the establishment of sex differences in brain and behaviour can be broadly divided into three stages. In the rodent, early life programming occurs during a perinatal sensitive period (the critical window) that is marked by the onset of gonadal steroid production in males. This occurs prenatally and around birth, a period during which there is no corresponding steroidogenesis in females. During this sensitive period, numerous neuroanatomical end points are differentiated in males versus females in a region-specific manner. In some areas, there are sex differences in apoptosis, in others, there are differences in synaptogenesis or neurogenesis. The juvenile hiatus begins shortly after birth and extends until puberty. It is characterized by low to non-existent steroid levels; however, during this time, social play behaviour is more frequent among males than among females, a behavioural sex difference that is organized during early life programming. The adult period is characterized by sex-specific hormonal milieus with males having high and steady levels of testosterone (with some daily and annual variations), whereas females undergo cyclic changes in hormone levels that are associated with ovulation. The steroid milieu of each sex activates the neural circuits of mating that were organized during the perinatal period. In this way, the gonadal phenotype and neural phenotype controlling mating behaviour are synchronized. Non-reproductive behaviours such as stress and anxiety, spatial learning, locomotion and social affiliation are also modulated by steroids in adulthood, and this modulation may or may not depend on earlier programming effects during the sensitive period.
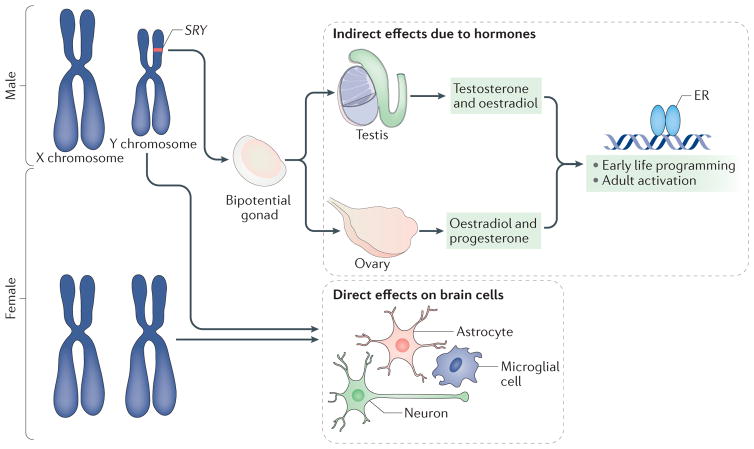
The differentiation of the bipotential gonad into a testis versus an ovary depends on the SRY gene, which is located on the Y chromosome. If present, the gonad will become a testis, and, if not, it will become an ovary. Testosterone and its aromatized end product oestradiol produced by the embryonic testis bind to their cognate receptors, which regulate gene transcription and exert an organizing influence on cells in the developing brain. Ovarian and testicular steroids also activate sex-specific physiology and behaviour in adulthood. Moreover, every cell in the brain is XY in male mammals and XX in female mammals. Genes on the X chromosome may be expressed at different levels in males and females (in cases in which they escape from X inactivation in females), whereas genes on the Y chromosome (such asSRY) are expressed only in the brain of males. In females, the presence of a second X chromosome could create a heterochromatic sink by monopolizing the cellular machinery used for epigenetic regulation and thus alter the expression of other genes in a manner that is distinct from males. Thus, the sex chromosomes can exert both specific and broad influences on the developing brain. ER, oestrogen receptor.
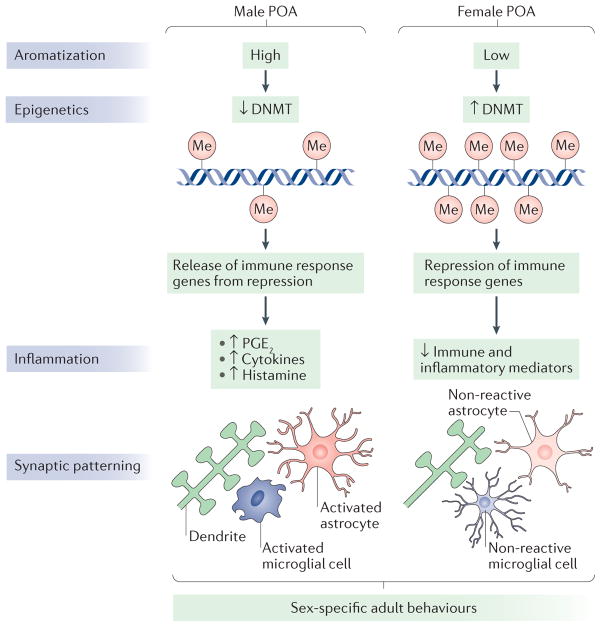
The rodent preoptic area (POA) is robustly sexually dimorphic and is therefore an ideal model system to understand the role of neuroepigenetics and neuroinflammation in sexual differentiation. The activity of DNA methyltransferase (DNMT) enzymes is higher in the developing POA of female rats than in that of males. This is because it is reduced in males, through unknown mechanisms, by the elevated oestradiol levels that are present owing to the aromatization of testosterone made in the testis. As a result, males have less DNA methylation (in the figure, indicated by ‘Me’) in POA cells. Indirect evidence suggests that the lower DNA methylation in males releases immune response genes from epigenetic repression, resulting in elevation of inflammatory signalling molecules such as prostaglandin E2 (PGE2), histamine and cytokines. PGE2 in particular is known to activate a signal transduction cascade that results in the formation of dendritic spine synapses on the dendrites of POA neurons. This process requires the participation of both astrocytes, as a putative source of glutamate, and neighbouring microglia, which provide much of the prostaglandin. In females, immune response genes are epigenetically silenced by higher levels of DNA methylation. PGE2 levels are therefore low, and both microglia and astrocytes are in a non-reactive state. As a result, females have half the density of dendritic spine synapses in the POA compared with males. These neuroanatomical sex differences undergird sex-specific reproductive behaviours, including male copulatory behaviour (both motivation to copulate and actual sexual performance), social scent marking, female oestrous cycling and maternal behaviour (for reviews, see REFS 44,114).
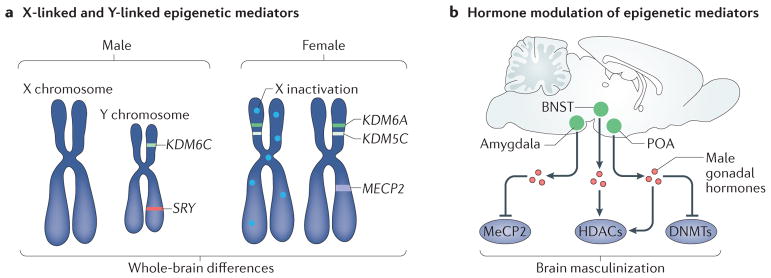
a | Several crucial epigenetic mediators are X chromosome and Y chromosome linked. These include the histone lysine demethylases KDM6A (also known as UTX) and KDM5C, which escape X inactivation and are thus expressed at higher levels in the female brain,. The Y-linked homologue of KDM6A, KDM6C (also known as UTY), is expressed at higher levels in the male brain. Sex-specific expression of epigenetic modifiers such as these has the potential to establish widespread sex differences in the chromatin landscape and gene expression and thus to drive structural and functional sex differences in the brain. X-linked chromatin-binding proteins, such as methyl CpG binding protein 2 (MeCP2), have also been shown to be important for establishment of brain sex differences. b | Several epigenetic mediators are regulated by sex specific gonadal hormones. For example, male gonadal hormones reduce the expression of MeCP2 in the amygdala, reduce DNA methyltransferase (DNMT) activity and methylation genome-wide in the preoptic area (POA), and alter methylation on specific promoters related to brain masculinization such as the oestrogen and progesterone receptors,. Hormonal modulation at the level of histone methylation and acetylation has also been demonstrated in the POA and bed nucleus of the stria terminalis (BNST), potentially mediating both active and repressive chromatin states,. HDACs, histone deacetylases.
References
-
- Griew S. Age and sex differences in probability learning of rats in a swimming T-maze. Gerontologia. 1968;14:197–203. - PubMed
-
- Maren S, De Oca B, Fanselow MS. Sex differences in hippocampal long-term potentiation (LTP) and Pavlovian fear conditioning in rats: positive correlation between LTP and contextual learning. Brain Res. 1994;661:25–34. - PubMed
-
- Perrot-Sinal TS. Sex differences in performance in the Morris water maze and the effects of initial nonstationary hidden platform training. Behav Neurosci. 1996;110:1309–1320. - PubMed
-
- Roof RL. Neonatal exogenous testosterone modifies sex difference in radial arm and Morris water maze perfromance in prebubescent and adult rats. Behav Brain Res. 1993;53:1–10. - PubMed
-
- Munoz-Cueto JA, Garcia-Segura LM, Ruiz-Marcos A. Developmental sex differences and effect of ovariectomy on the number of cortical pyramidal cell dendritic spines. Brain Res. 1990;515:64–68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
