

Phenotypic Plasticity and Cell Fate Decisions in Cancer: Insights from Dynamical Systems Theory
- PMID: 28640191
- PMCID: PMC5532606
- DOI: 10.3390/cancers9070070
Phenotypic Plasticity and Cell Fate Decisions in Cancer: Insights from Dynamical Systems Theory
Abstract
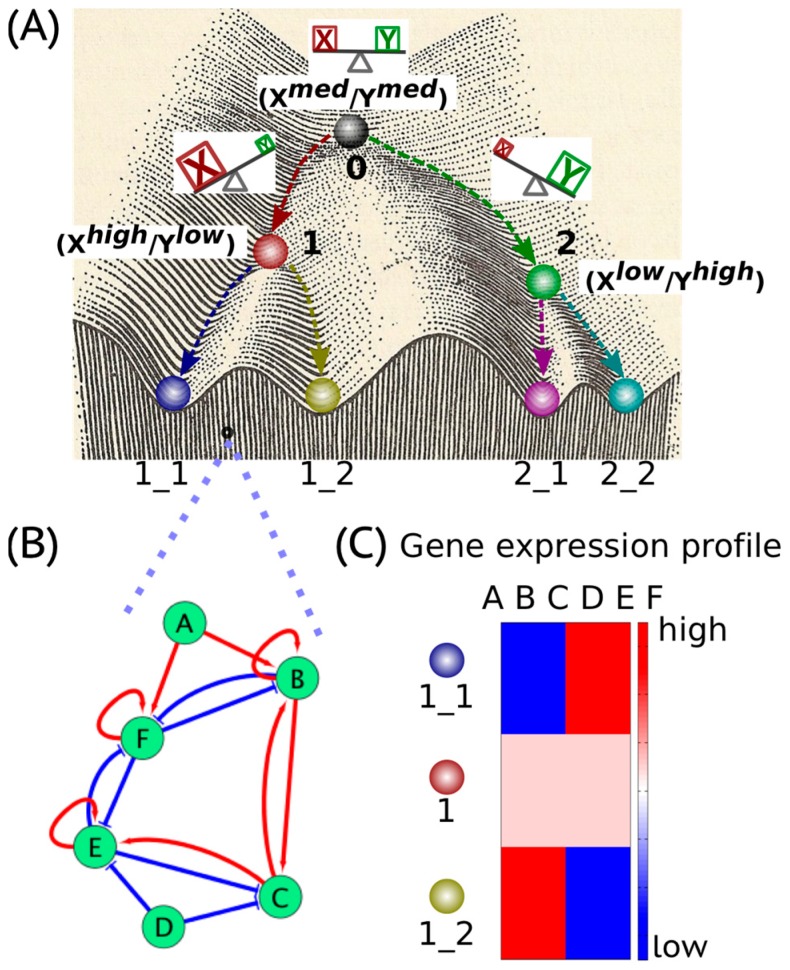
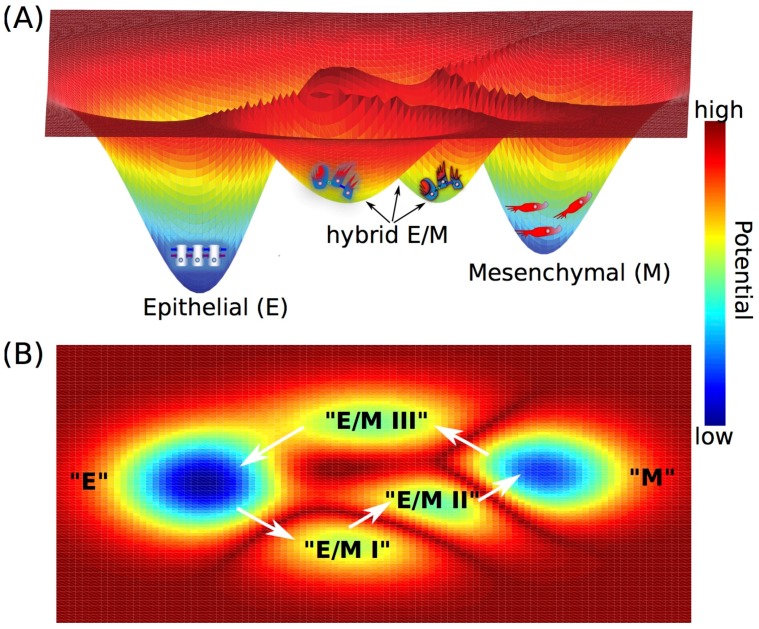
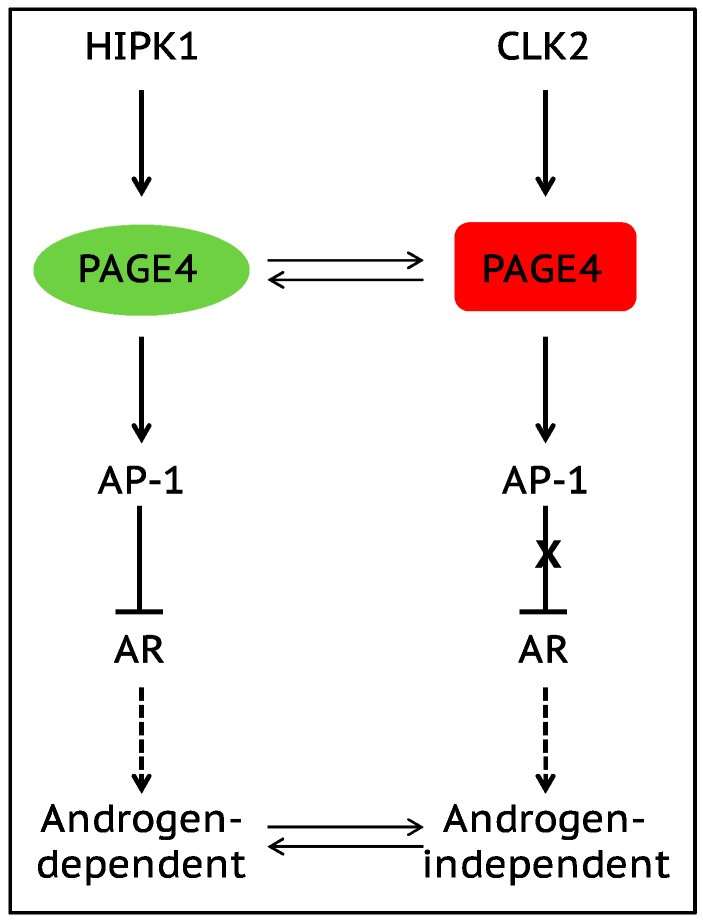
Waddington's epigenetic landscape, a famous metaphor in developmental biology, depicts how a stem cell progresses from an undifferentiated phenotype to a differentiated one. The concept of "landscape" in the context of dynamical systems theory represents a high-dimensional space, in which each cell phenotype is considered as an "attractor" that is determined by interactions between multiple molecular players, and is buffered against environmental fluctuations. In addition, biological noise is thought to play an important role during these cell-fate decisions and in fact controls transitions between different phenotypes. Here, we discuss the phenotypic transitions in cancer from a dynamical systems perspective and invoke the concept of "cancer attractors"-hidden stable states of the underlying regulatory network that are not occupied by normal cells. Phenotypic transitions in cancer occur at varying levels depending on the context. Using epithelial-to-mesenchymal transition (EMT), cancer stem-like properties, metabolic reprogramming and the emergence of therapy resistance as examples, we illustrate how phenotypic plasticity in cancer cells enables them to acquire hybrid phenotypes (such as hybrid epithelial/mesenchymal and hybrid metabolic phenotypes) that tend to be more aggressive and notoriously resilient to therapies such as chemotherapy and androgen-deprivation therapy. Furthermore, we highlight multiple factors that may give rise to phenotypic plasticity in cancer cells, such as (a) multi-stability or oscillatory behaviors governed by underlying regulatory networks involved in cell-fate decisions in cancer cells, and (b) network rewiring due to conformational dynamics of intrinsically disordered proteins (IDPs) that are highly enriched in cancer cells. We conclude by discussing why a therapeutic approach that promotes "recanalization", i.e., the exit from "cancer attractors" and re-entry into "normal attractors", is more likely to succeed rather than a conventional approach that targets individual molecules/pathways.
Keywords: EMT; cancer attractors; cell fate decision; gene network dynamics; intrinsically disordered proteins; therapy resistance.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Waddington C.H. The Strategy of the Genes. George Allen & Unwin; London, UK: 1957.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
