

Ribosomal DNA copy number loss and sequence variation in cancer
- PMID: 28640831
- PMCID: PMC5480814
- DOI: 10.1371/journal.pgen.1006771
Ribosomal DNA copy number loss and sequence variation in cancer
Abstract
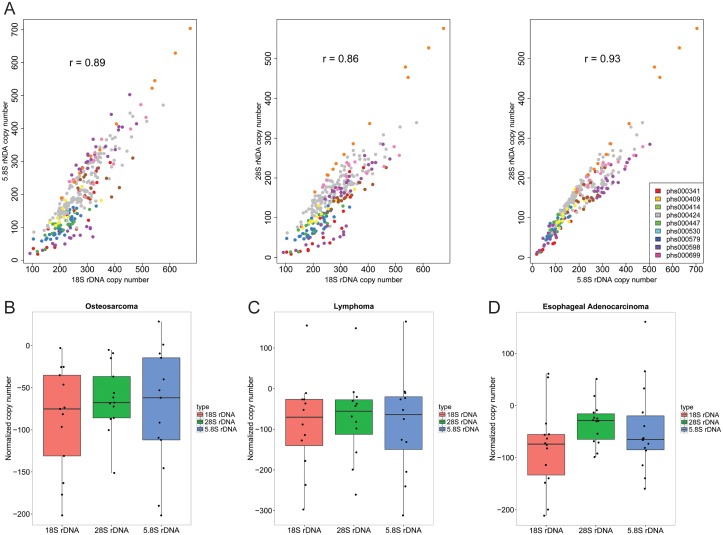
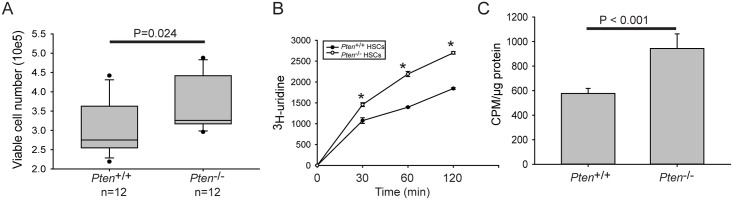
Ribosomal DNA is one of the most variable regions in the human genome with respect to copy number. Despite the importance of rDNA for cellular function, we know virtually nothing about what governs its copy number, stability, and sequence in the mammalian genome due to challenges associated with mapping and analysis. We applied computational and droplet digital PCR approaches to measure rDNA copy number in normal and cancer states in human and mouse genomes. We find that copy number and sequence can change in cancer genomes. Counterintuitively, human cancer genomes show a loss of copies, accompanied by global copy number co-variation. The sequence can also be more variable in the cancer genome. Cancer genomes with lower copies have mutational evidence of mTOR hyperactivity. The PTEN phosphatase is a tumor suppressor that is critical for genome stability and a negative regulator of the mTOR kinase pathway. Surprisingly, but consistent with the human cancer genomes, hematopoietic cancer stem cells from a Pten-/- mouse model for leukemia have lower rDNA copy number than normal tissue, despite increased proliferation, rRNA production, and protein synthesis. Loss of copies occurs early and is associated with hypersensitivity to DNA damage. Therefore, copy loss is a recurrent feature in cancers associated with mTOR activation. Ribosomal DNA copy number may be a simple and useful indicator of whether a cancer will be sensitive to DNA damaging treatments.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- McStay B (2016) Nucleolar organizer regions: genomic 'dark matter' requiring illumination. Genes Dev 30: 1598–1610. doi: 10.1101/gad.283838.116 - DOI - PMC - PubMed
-
- Forsburg SL (2013) The CINs of the centromere. Biochem Soc Trans 41: 1706–1711. doi: 10.1042/BST20130146 - DOI - PMC - PubMed
-
- De Lange T (2005) Telomere-related genome instability in cancer. Cold Spring Harb Symp Quant Biol 70: 197–204. doi: 10.1101/sqb.2005.70.032 - DOI - PubMed
-
- Gibbons JG, Branco AT, Godinho SA, Yu S, Lemos B (2015) Concerted copy number variation balances ribosomal DNA dosage in human and mouse genomes. Proc Natl Acad Sci U S A 112: 2485–2490. doi: 10.1073/pnas.1416878112 - DOI - PMC - PubMed
-
- Long EO, Dawid IB (1980) Repeated genes in eukaryotes. Annu Rev Biochem 49: 727–764. doi: 10.1146/annurev.bi.49.070180.003455 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
