

The interaction between progranulin and prosaposin is mediated by granulins and the linker region between saposin B and C
- PMID: 28640985
- PMCID: PMC5630500
- DOI: 10.1111/jnc.14110
The interaction between progranulin and prosaposin is mediated by granulins and the linker region between saposin B and C
Abstract
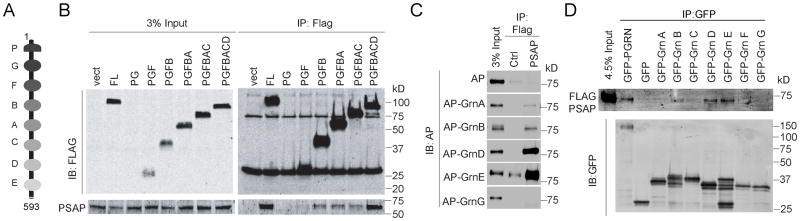
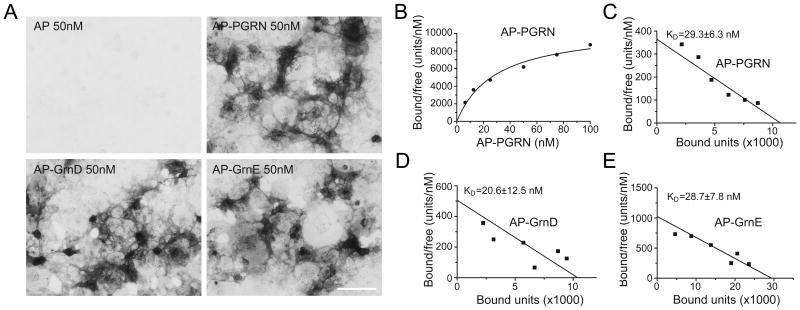
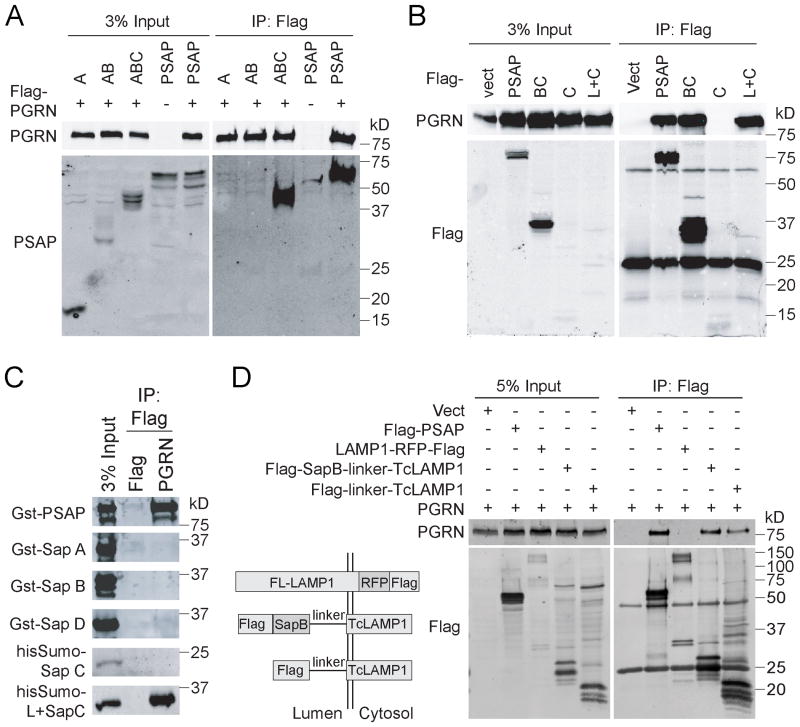
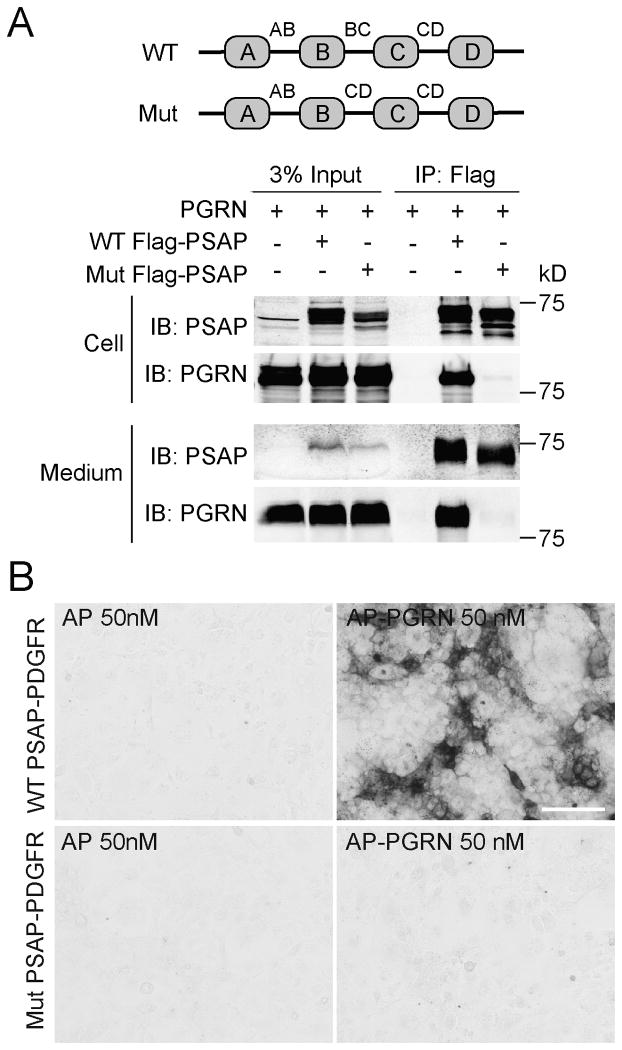
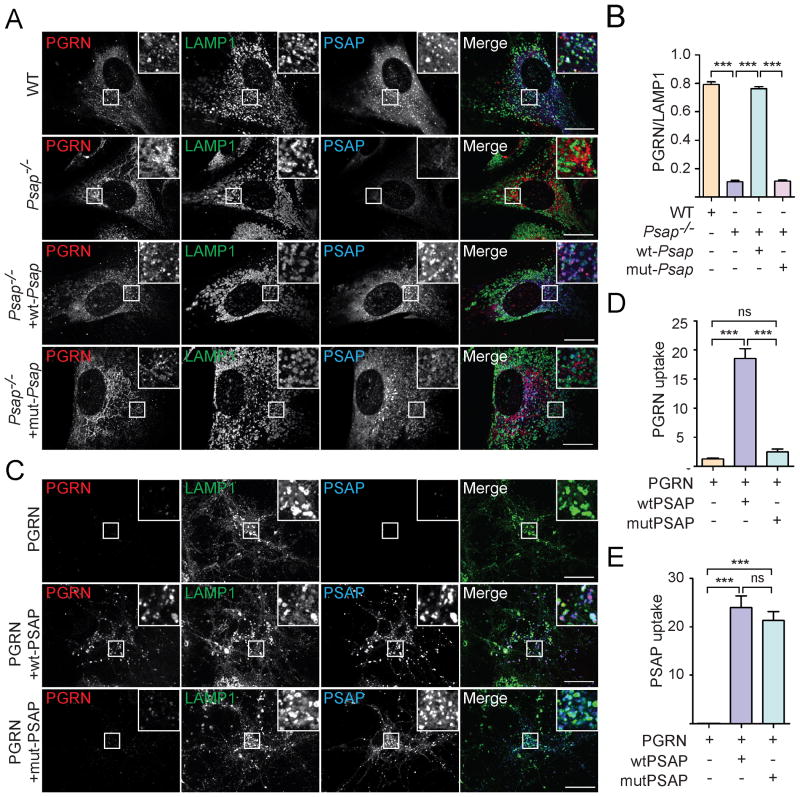
The frontotemporal lobar degeneration (FTLD) protein progranulin (PGRN) is essential for proper lysosomal function. PGRN localizes in the lysosomal compartment within the cell. Prosaposin (PSAP), the precursor of lysosomal saposin activators (saposin A, B, C, D), physically interacts with PGRN. Previously, we have shown that PGRN and PSAP facilitate each other's lysosomal trafficking. Here, we report that the interaction between PSAP and PGRN requires the linker region of saposin B and C (BC linker). PSAP protein with the BC linker mutated, fails to interact with PGRN and deliver PGRN to lysosomes in the biosynthetic and endocytic pathways. On the other hand, PGRN interacts with PSAP through multiple granulin motifs. Granulin D and E bind to PSAP with similar affinity as full-length PGRN. Read the Editorial Comment for this article on page 154.
Keywords: frontotemporal lobar degeneration; lysosomal storage diseases; lysosome; neuronal ceroid lipofuscinosis; progranulin; prosaposin.
© 2017 International Society for Neurochemistry.
Conflict of interest statement
Figures
Comment in
-
Another piece in the progranulin puzzle: special binding between progranulin and prosaposin creates additional lysosomal access: An Editorial Comment for 'The interaction between progranulin and prosaposin is mediated by granulins and the linker region between saposin B and C' on page 236.J Neurochem. 2017 Oct;143(2):154-157. doi: 10.1111/jnc.14125. Epub 2017 Aug 4. J Neurochem. 2017. PMID: 28776681 Free PMC article.
References
-
- Almeida MR, Macario MC, Ramos L, Baldeiras I, Ribeiro MH, Santana I. Portuguese family with the co-occurrence of frontotemporal lobar degeneration and neuronal ceroid lipofuscinosis phenotypes due to progranulin gene mutation. Neurobiol Aging 2016 - PubMed
-
- Arai T, Hasegawa M, Akiyama H, Ikeda K, Nonaka T, Mori H, Mann D, Tsuchiya K, Yoshida M, Hashizume Y, et al. TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem Biophys Res Commun. 2006;351:602–611. - PubMed
-
- Baker M, Mackenzie IR, Pickering-Brown SM, Gass J, Rademakers R, Lindholm C, Snowden J, Adamson J, Sadovnick AD, Rollinson S, et al. Mutations in progranulin cause tau-negative frontotemporal dementia linked to chromosome 17. Nature. 2006;442:916–919. - PubMed
-
- Bateman A, Bennett HP. The granulin gene family: from cancer to dementia. Bioessays. 2009;31:1245–1254. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
