

Investigation of protein synthesis in Drosophila larvae using puromycin labelling
- PMID: 28642244
- PMCID: PMC5576084
- DOI: 10.1242/bio.026294
Investigation of protein synthesis in Drosophila larvae using puromycin labelling
Abstract
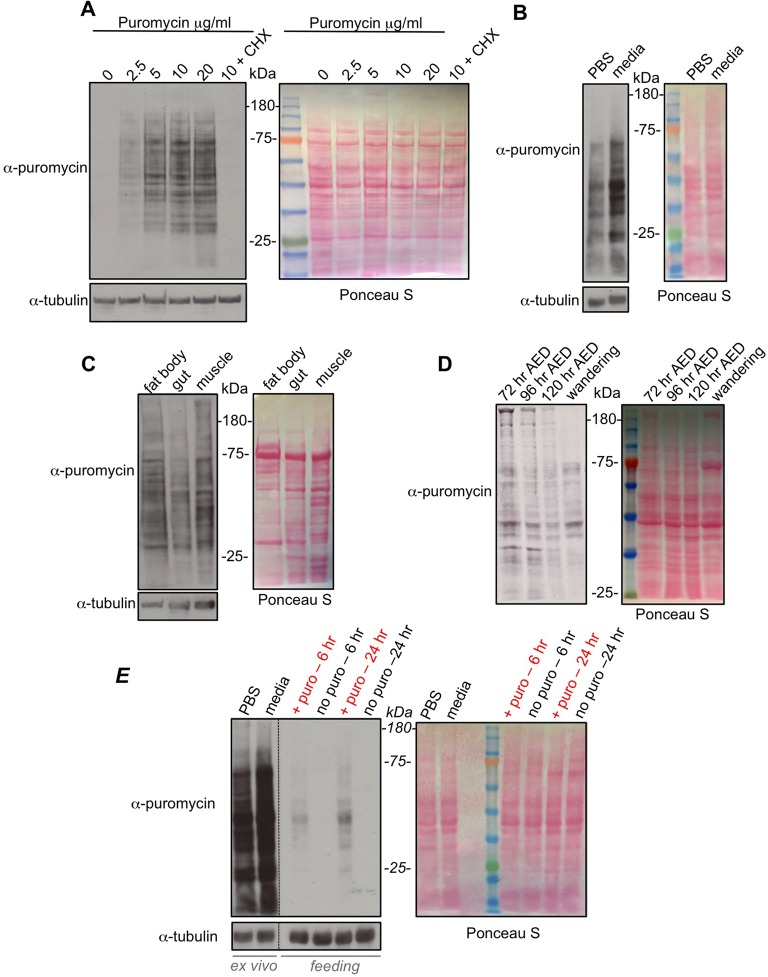
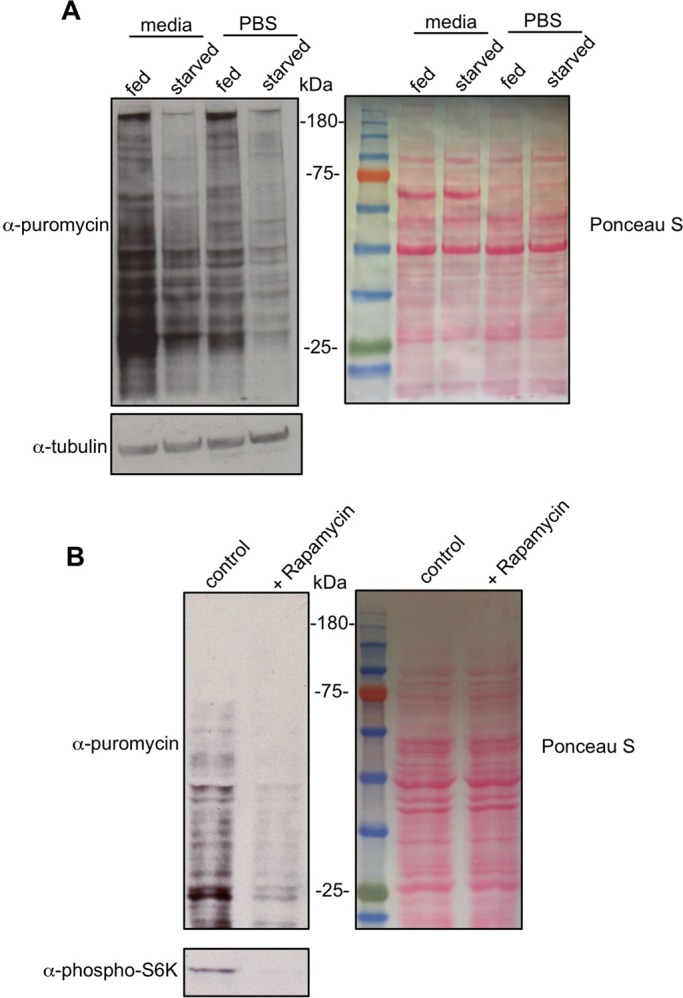
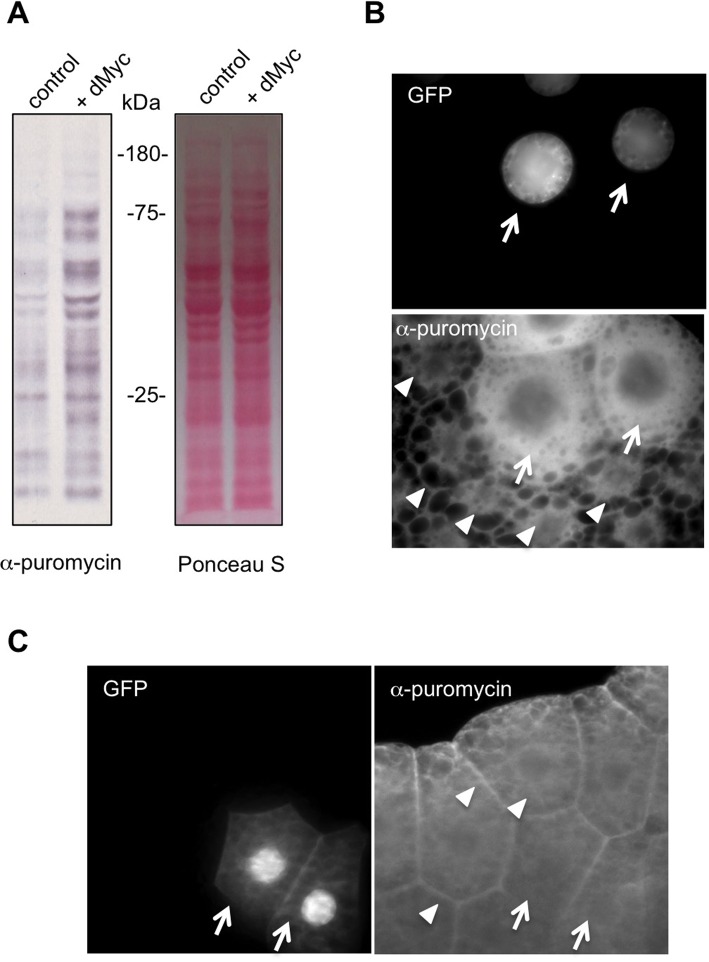
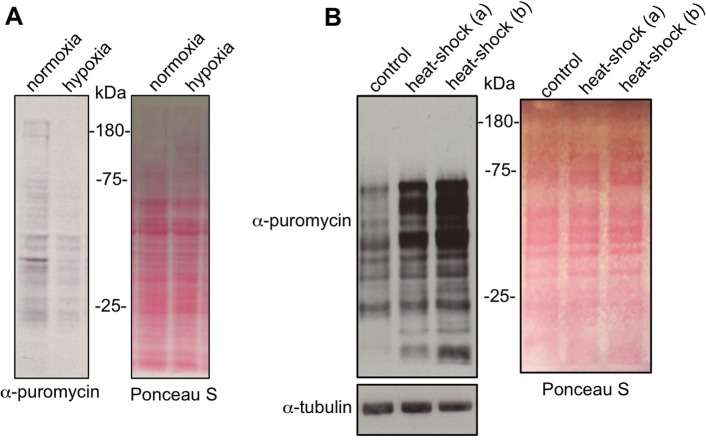
Translational control of gene expression is an important regulator of growth, homeostasis and aging in Drosophila The ability to measure changes in protein synthesis in response to genetic and environmental cues is therefore important in studying these processes. Here we describe a simple and cost-effective approach to assay protein synthesis in Drosophila larval cells and tissues. The method is based on the incorporation of puromycin into nascent peptide chains. Using an ex vivo approach, we label newly synthesized peptides in larvae with puromycin and then measure levels of new protein synthesis using an anti-puromycin antibody. We show that this method can detect changes in protein synthesis in specific cells and tissues in the larvae, either by immunostaining or western blotting. We find that the assay reliably detects changes in protein synthesis induced by two known stimulators of mRNA translation - the nutrient/TORC1 kinase pathway and the transcription factor dMyc. We also use the assay to describe how protein synthesis changes through larval development and in response to two environmental stressors - hypoxia and heat shock. We propose that this puromycin-labelling assay is a simple but robust method to detect protein synthesis changes at the levels of cells, tissues or whole body in Drosophila.
Keywords: Drosophila; Heat shock; Hypoxia; Nutrients; Protein synthesis; TOR kinase; dMyc; mRNA translation.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Cook D., Nuro E., Jones E. V., Altimimi H. F., Farmer W. T., Gandin V., Hanna E., Zong R., Barbon A., Nelson D. L. et al. (2014). FXR1P limits long-term memory, long-lasting synaptic potentiation, and de novo GluA2 translation. Cell Rep. 9, 1402-1416. 10.1016/j.celrep.2014.10.028 - DOI - PMC - PubMed
-
- Dalet A., Argüello R. J., Combes A., Spinelli L., Jaeger S., Fallet M., Vu Manh T. P., Mendes A., Perego J., Reverendo M. et al. (2017). Protein synthesis inhibition and GADD34 control IFN-beta heterogeneous expression in response to dsRNA. EMBO J. 36, 761-782. 10.15252/embj.201695000 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
