

Modulation of Endoplasmic Reticulum Stress Controls CD4+ T-cell Activation and Antitumor Function
- PMID: 28642246
- PMCID: PMC5585019
- DOI: 10.1158/2326-6066.CIR-17-0081
Modulation of Endoplasmic Reticulum Stress Controls CD4+ T-cell Activation and Antitumor Function
Abstract
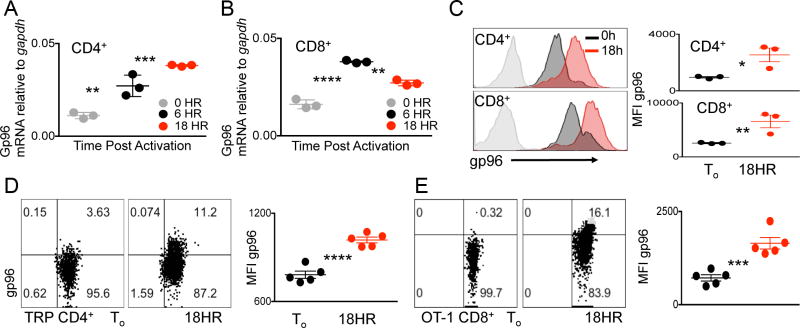
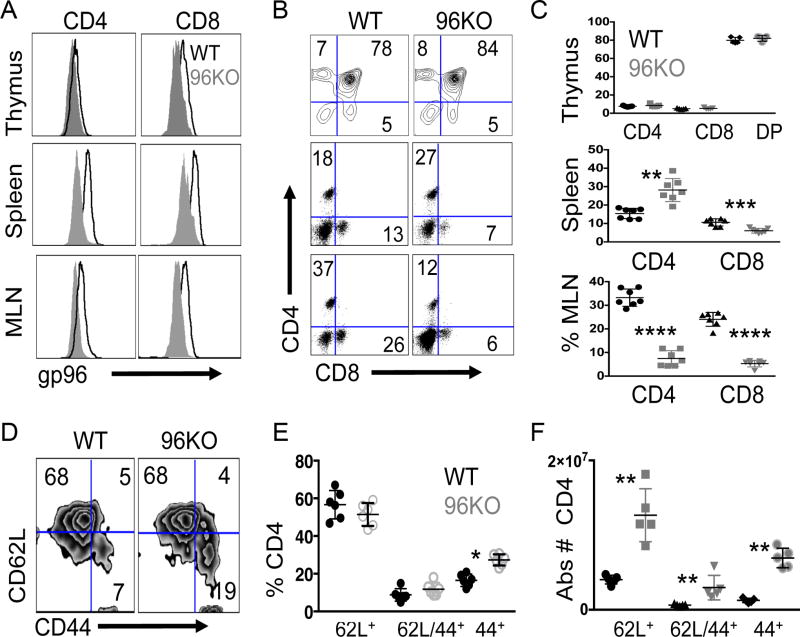
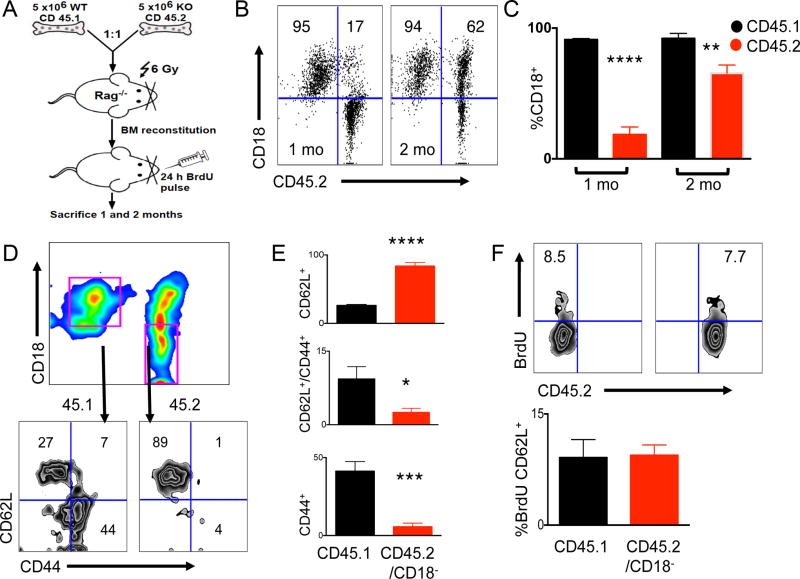
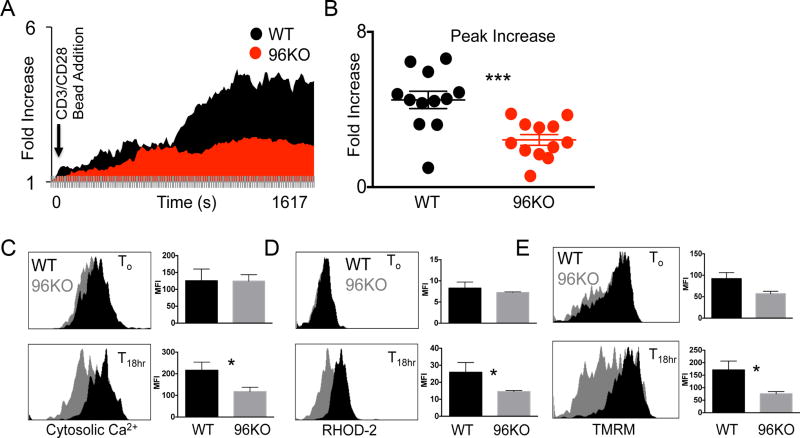
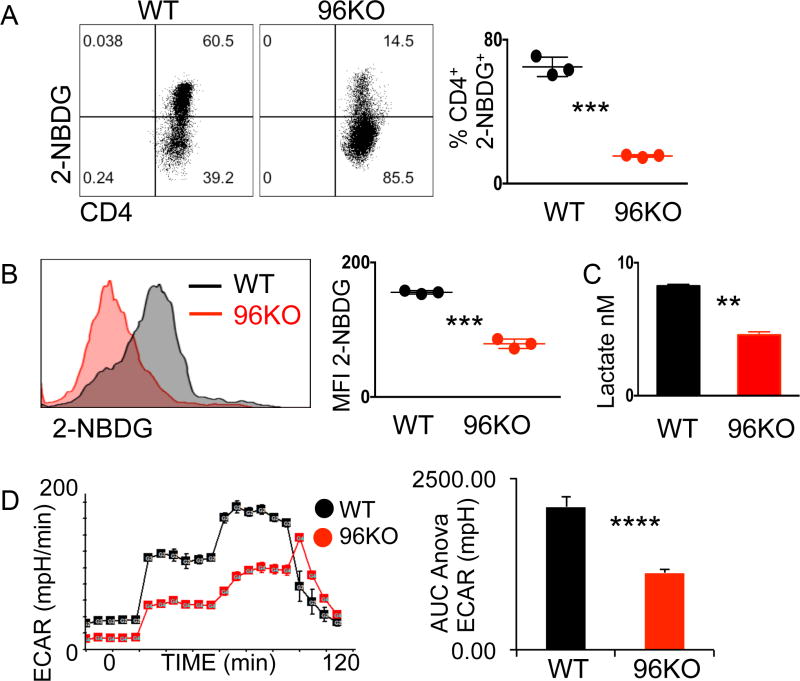
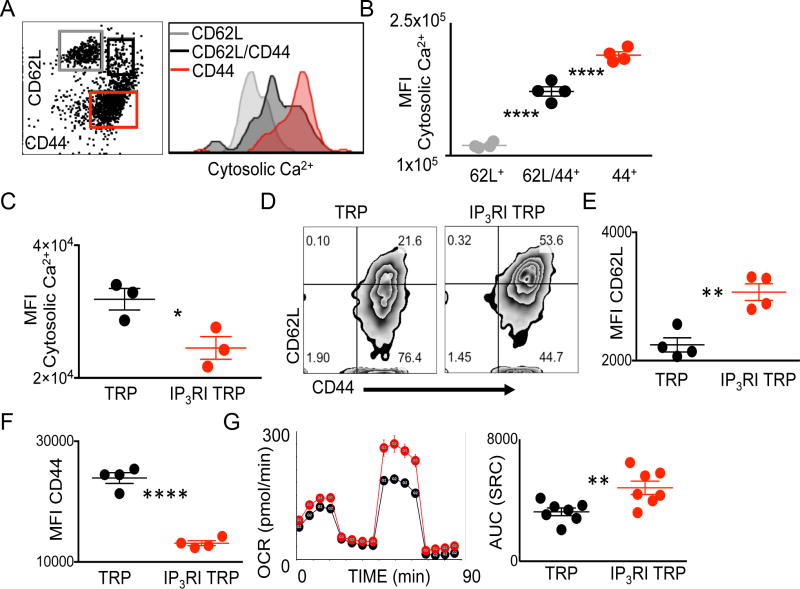
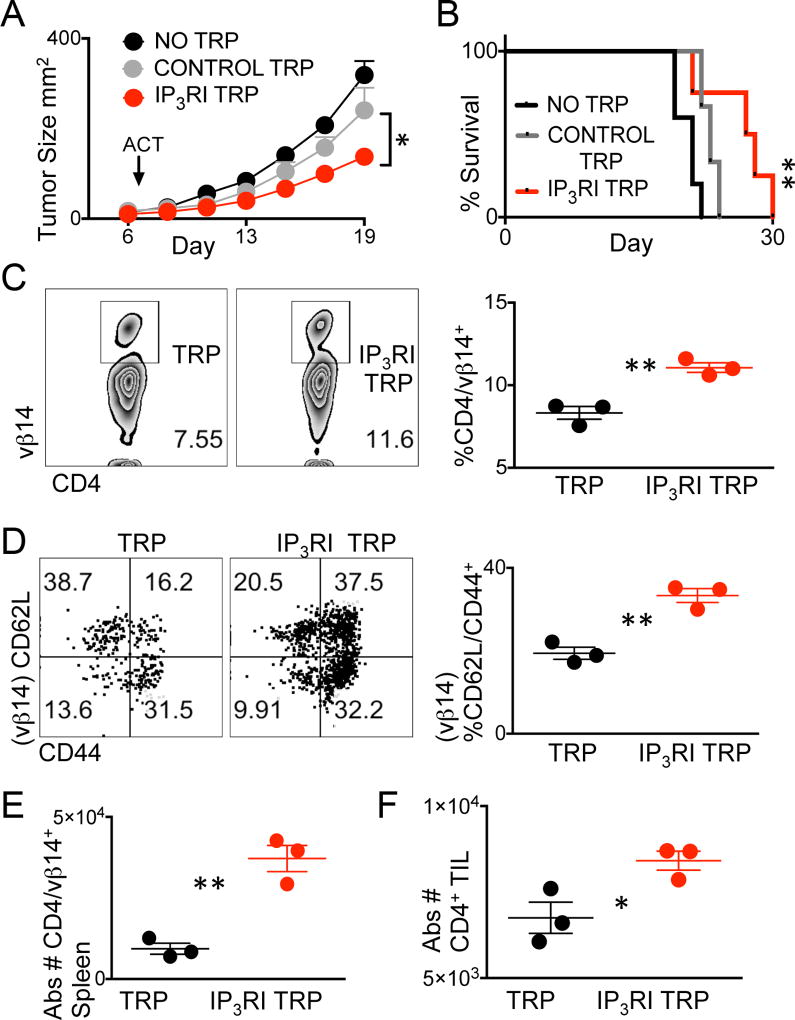
The endoplasmic reticulum (ER) is an energy-sensing organelle with intimate ties to programming cell activation and metabolic fate. T-cell receptor (TCR) activation represents a form of acute cell stress and induces mobilization of ER Ca2+ stores. The role of the ER in programming T-cell activation and metabolic fate remains largely undefined. Gp96 is an ER protein with functions as a molecular chaperone and Ca2+ buffering protein. We hypothesized that the ER stress response may be important for CD4+ T-cell activation and that gp96 may be integral to this process. To test our hypothesis, we utilized genetic deletion of the gp96 gene Hsp90b1 in a CD4+ T cell-specific manner. We show that gp96-deficient CD4+ T cells cannot undergo activation-induced glycolysis due to defective Ca2+ mobilization upon TCR engagement. We found that activating naïve CD4+ T cells while inhibiting ER Ca2+ exchange, through pharmacological blockade of the ER Ca2+ channel inositol trisphosphate receptor (IP3R), led to a reduction in cytosolic Ca2+ content and generated a pool of CD62Lhigh/CD44low CD4+ T cells compared with wild-type (WT) matched controls. In vivo IP3R-inhibited CD4+ T cells exhibited elevated tumor control above WT T cells. Together, these data show that ER-modulated cytosolic Ca2+ plays a role in defining CD4+ T-cell phenotype and function. Factors associated with the ER stress response are suitable targets for T cell-based immunotherapies. Cancer Immunol Res; 5(8); 666-75. ©2017 AACR.
©2017 American Association for Cancer Research.
Conflict of interest statement
Figures
Similar articles
-
Disruption of calcium transfer from ER to mitochondria links alterations of mitochondria-associated ER membrane integrity to hepatic insulin resistance.Diabetologia. 2016 Mar;59(3):614-23. doi: 10.1007/s00125-015-3829-8. Epub 2015 Dec 10. Diabetologia. 2016. PMID: 26660890
-
Inositol 1,4,5-trisphosphate receptor-isoform diversity in cell death and survival.Biochim Biophys Acta. 2014 Oct;1843(10):2164-83. doi: 10.1016/j.bbamcr.2014.03.007. Epub 2014 Mar 15. Biochim Biophys Acta. 2014. PMID: 24642269 Review.
-
Tespa1 is a novel component of mitochondria-associated endoplasmic reticulum membranes and affects mitochondrial calcium flux.Biochem Biophys Res Commun. 2013 Apr 12;433(3):322-6. doi: 10.1016/j.bbrc.2013.02.099. Epub 2013 Mar 15. Biochem Biophys Res Commun. 2013. PMID: 23501103
-
Inherent ER stress in pancreatic islet β cells causes self-recognition by autoreactive T cells in type 1 diabetes.J Autoimmun. 2016 Aug;72:33-46. doi: 10.1016/j.jaut.2016.04.009. Epub 2016 May 9. J Autoimmun. 2016. PMID: 27173406 Free PMC article.
-
Regulation of inositol 1,4,5-trisphosphate receptors during endoplasmic reticulum stress.Biochim Biophys Acta. 2013 Jul;1833(7):1612-24. doi: 10.1016/j.bbamcr.2013.01.026. Epub 2013 Feb 1. Biochim Biophys Acta. 2013. PMID: 23380704 Review.
Cited by
-
XBP1 is required in Th2 polarization induction in airway allergy.Theranostics. 2022 Jul 11;12(12):5337-5349. doi: 10.7150/thno.75100. eCollection 2022. Theranostics. 2022. PMID: 35910793 Free PMC article.
-
Endoplasmic Reticulum Stress Contributes to Mitochondrial Exhaustion of CD8+ T Cells.Cancer Immunol Res. 2019 Mar;7(3):476-486. doi: 10.1158/2326-6066.CIR-18-0182. Epub 2019 Jan 18. Cancer Immunol Res. 2019. PMID: 30659052 Free PMC article.
-
Endoplasmic Reticulum Stress in Tuberculosis: Molecular Bases and Pathophysiological Implications in the Immunopathogenesis of the Disease.Int J Mol Sci. 2025 May 9;26(10):4522. doi: 10.3390/ijms26104522. Int J Mol Sci. 2025. PMID: 40429667 Free PMC article. Review.
-
The Unfolded Protein Response at the Tumor-Immune Interface.Front Immunol. 2022 Feb 14;13:823157. doi: 10.3389/fimmu.2022.823157. eCollection 2022. Front Immunol. 2022. PMID: 35237269 Free PMC article. Review.
-
Roles of Endoplasmic Reticulum Stress in Immune Responses.Mol Cells. 2018 Aug 31;41(8):705-716. doi: 10.14348/molcells.2018.0241. Epub 2018 Jul 30. Mol Cells. 2018. PMID: 30078231 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
