

Nonribosomal peptide synthetase biosynthetic clusters of ESKAPE pathogens
- PMID: 28642945
- PMCID: PMC5551671
- DOI: 10.1039/c7np00029d
Nonribosomal peptide synthetase biosynthetic clusters of ESKAPE pathogens
Abstract
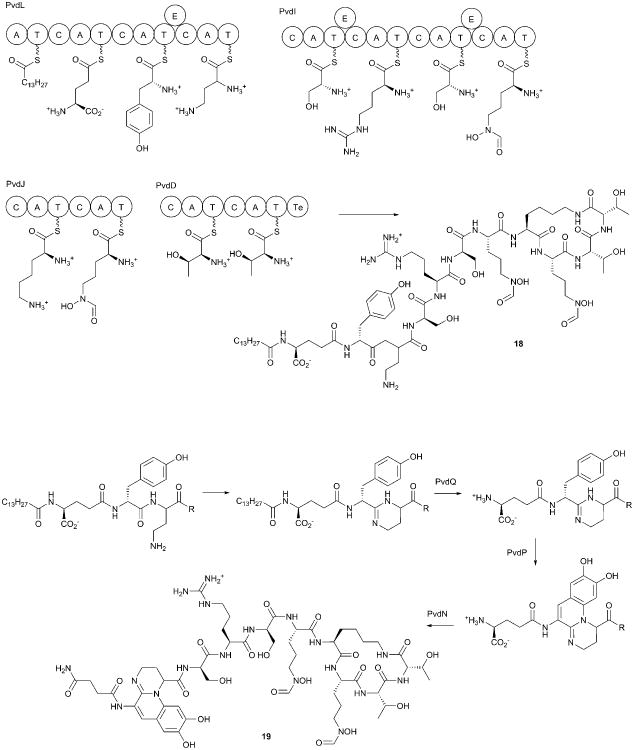
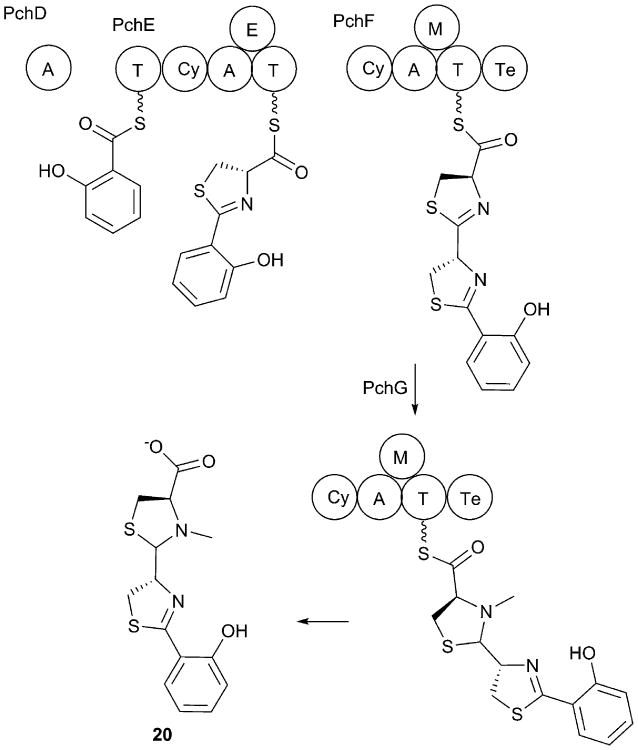
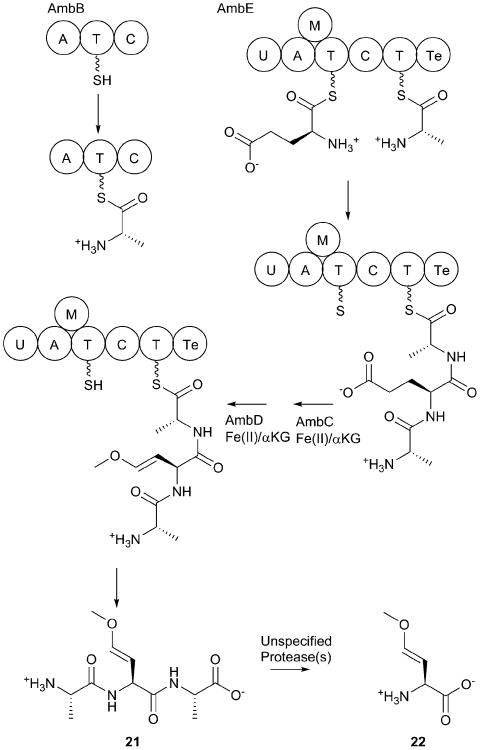
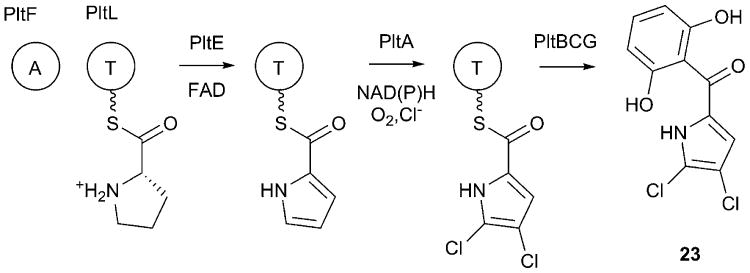
Covering: up to 2017.Natural products are important secondary metabolites produced by bacterial and fungal species that play important roles in cellular growth and signaling, nutrient acquisition, intra- and interspecies communication, and virulence. A subset of natural products is produced by nonribosomal peptide synthetases (NRPSs), a family of large, modular enzymes that function in an assembly line fashion. Because of the pharmaceutical activity of many NRPS products, much effort has gone into the exploration of their biosynthetic pathways and the diverse products they make. Many interesting NRPS pathways have been identified and characterized from both terrestrial and marine bacterial sources. Recently, several NRPS pathways in human commensal bacterial species have been identified that produce molecules with antibiotic activity, suggesting another source of interesting NRPS pathways may be the commensal and pathogenic bacteria that live on the human body. The ESKAPE pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp.) have been identified as a significant cause of human bacterial infections that are frequently multidrug resistant. The emerging resistance profile of these organisms has prompted calls from multiple international agencies to identify novel antibacterial targets and develop new approaches to treat infections from ESKAPE pathogens. Each of these species contains several NRPS biosynthetic gene clusters. While some have been well characterized and produce known natural products with important biological roles in microbial physiology, others have yet to be investigated. This review catalogs the NRPS pathways of ESKAPE pathogens. The exploration of novel NRPS products may lead to a better understanding of the chemical communication used by human pathogens and potentially to the discovery of novel therapeutic approaches.
Figures
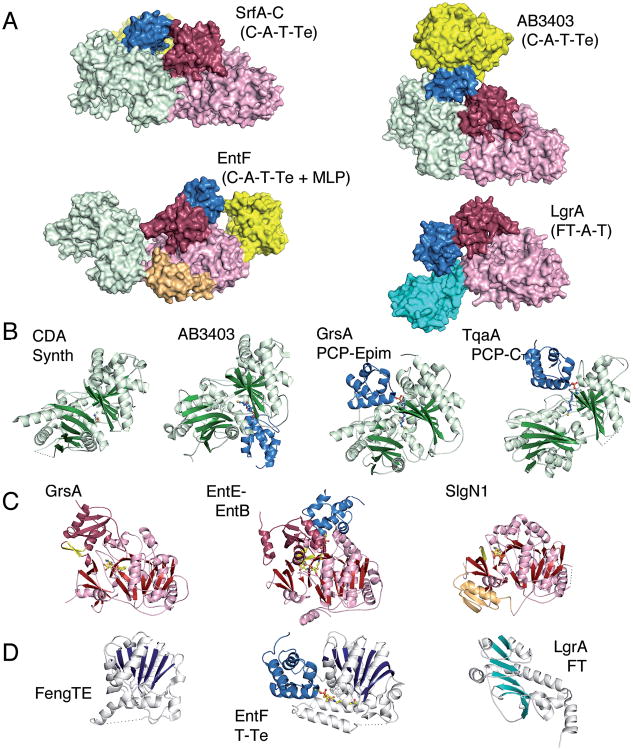
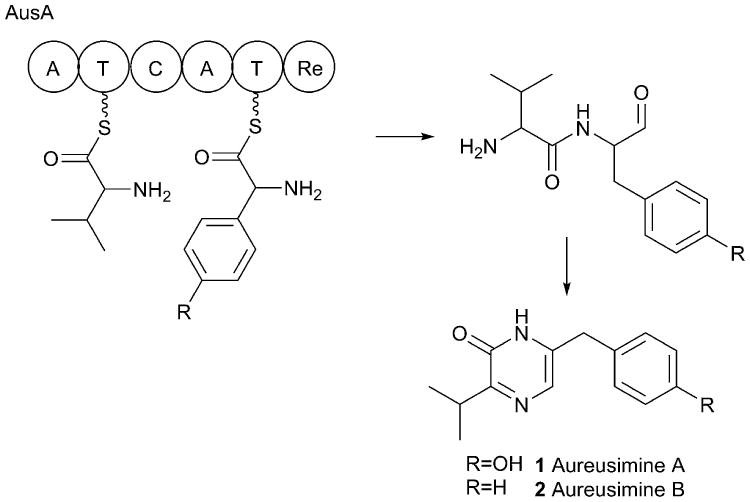
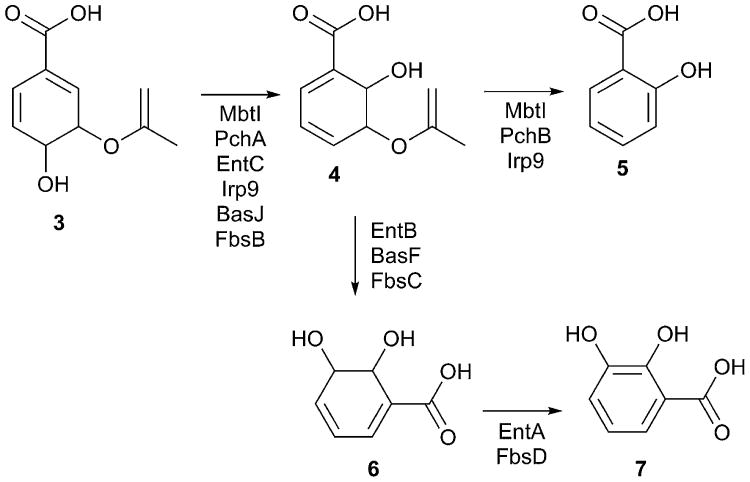
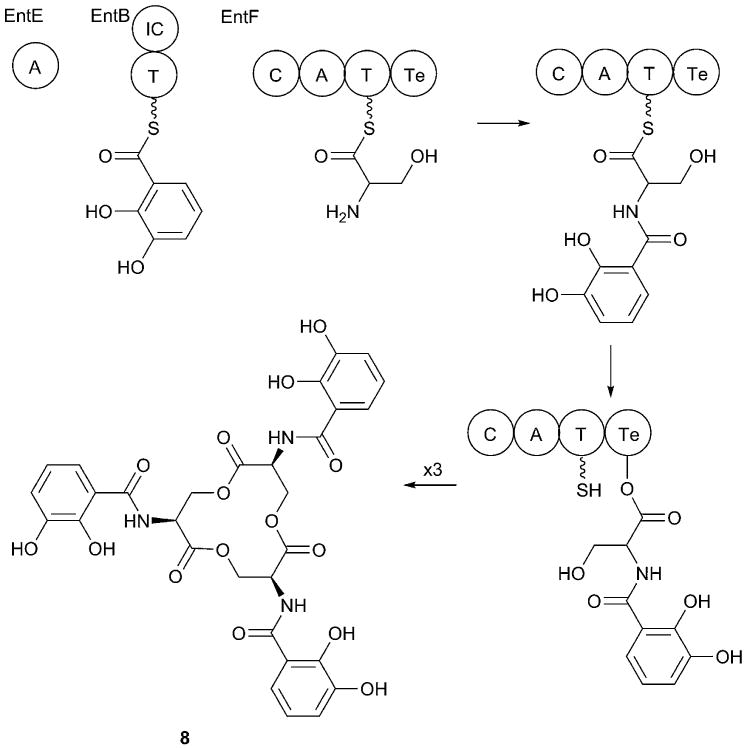
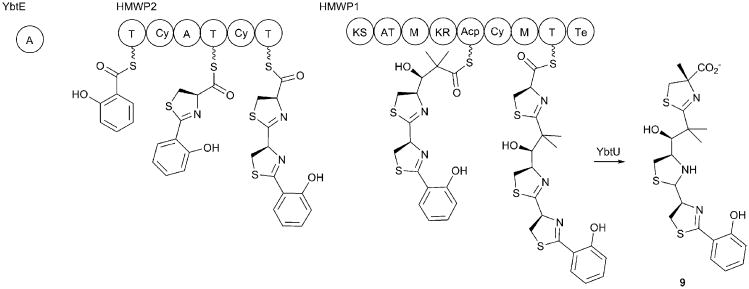
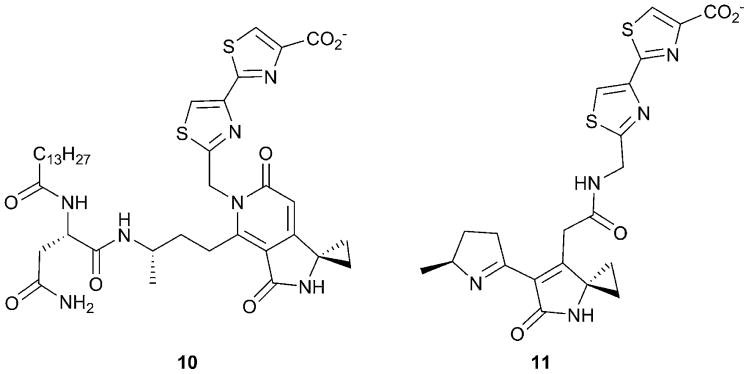
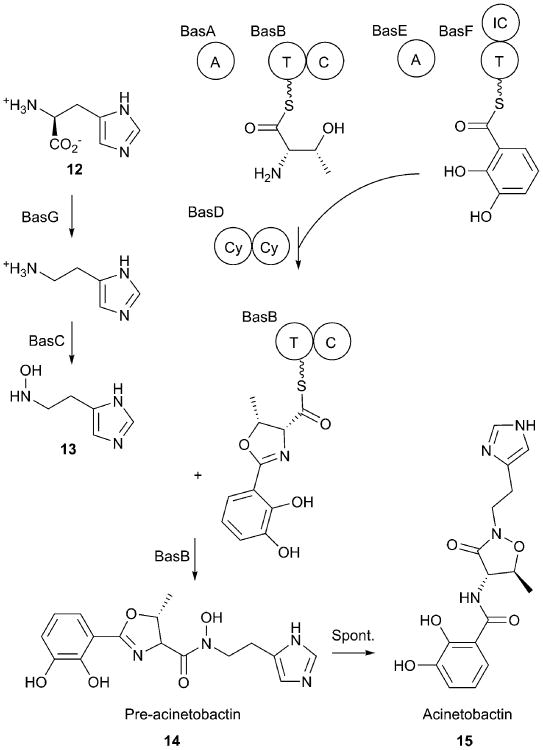
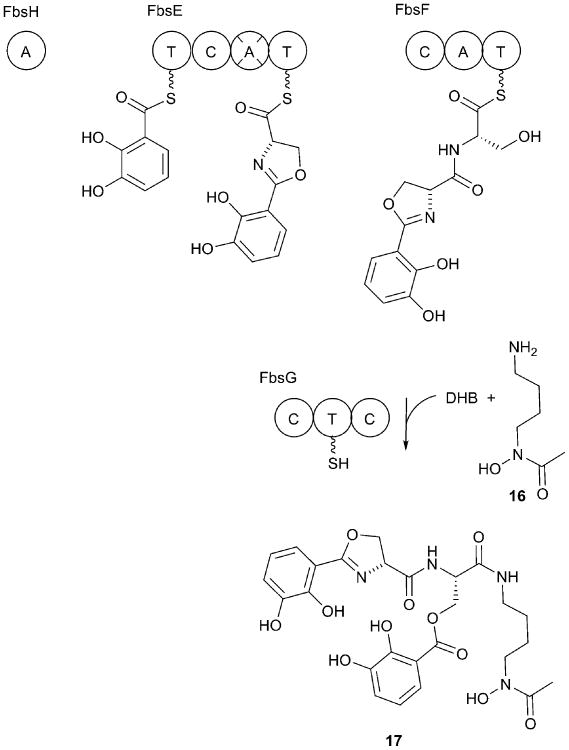
References
-
- Gilhuus-Moe CC, Kristensen T, Bredesen JE, Zimmer T, Laland SG. FEBS Lett. 1970;7:287–290. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous