

IRF6 expression in basal epithelium partially rescues Irf6 knockout mice
- PMID: 28643456
- PMCID: PMC5552062
- DOI: 10.1002/dvdy.24537
IRF6 expression in basal epithelium partially rescues Irf6 knockout mice
Abstract
Background: Mutations in IRF6, CHUK (IKKA), and RIPK4 can lead to a disease spectrum that includes cutaneous, limb, and craniofacial malformations. Loss of these alleles in the mouse leads to perinatal lethality and severe cutaneous, limb, and craniofacial defects also. Genetic rescue in the mouse has been shown for Ikka and Ripk4.
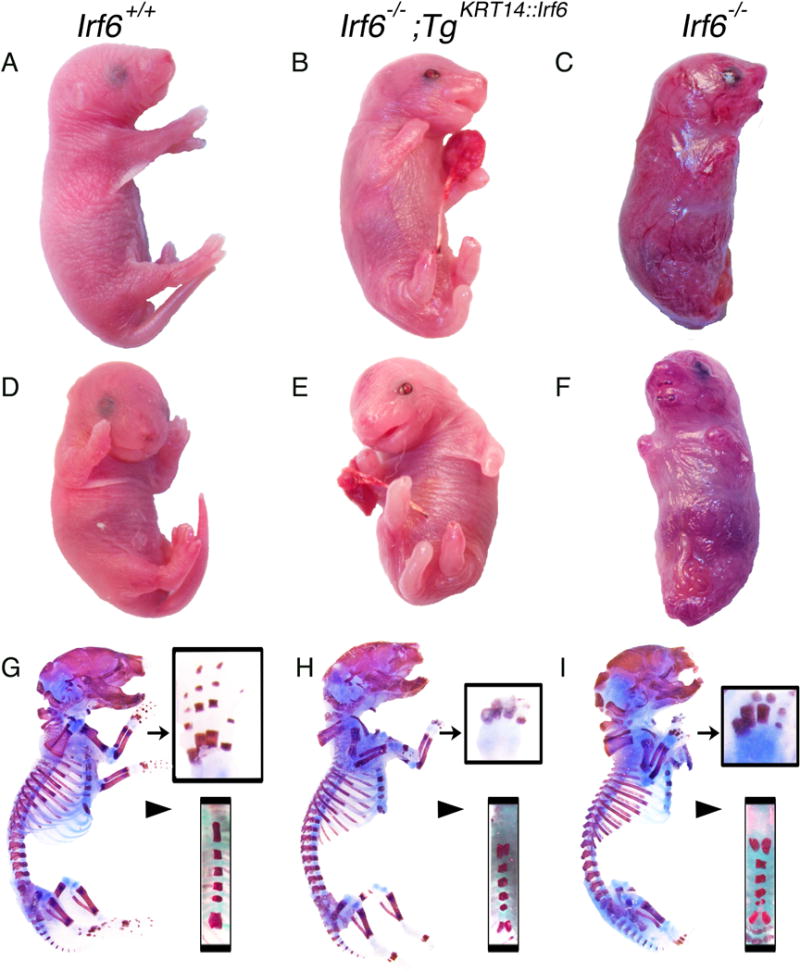
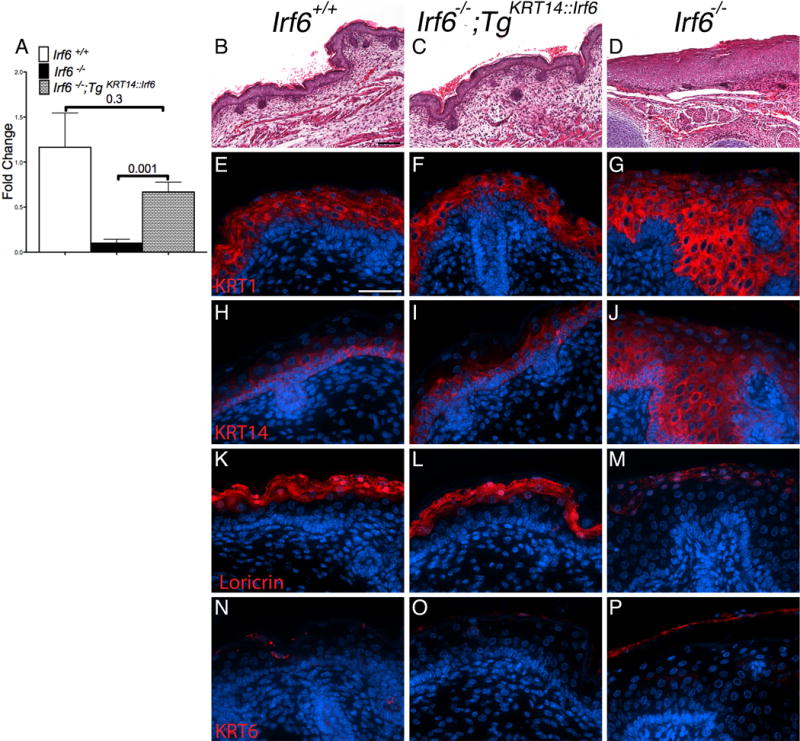
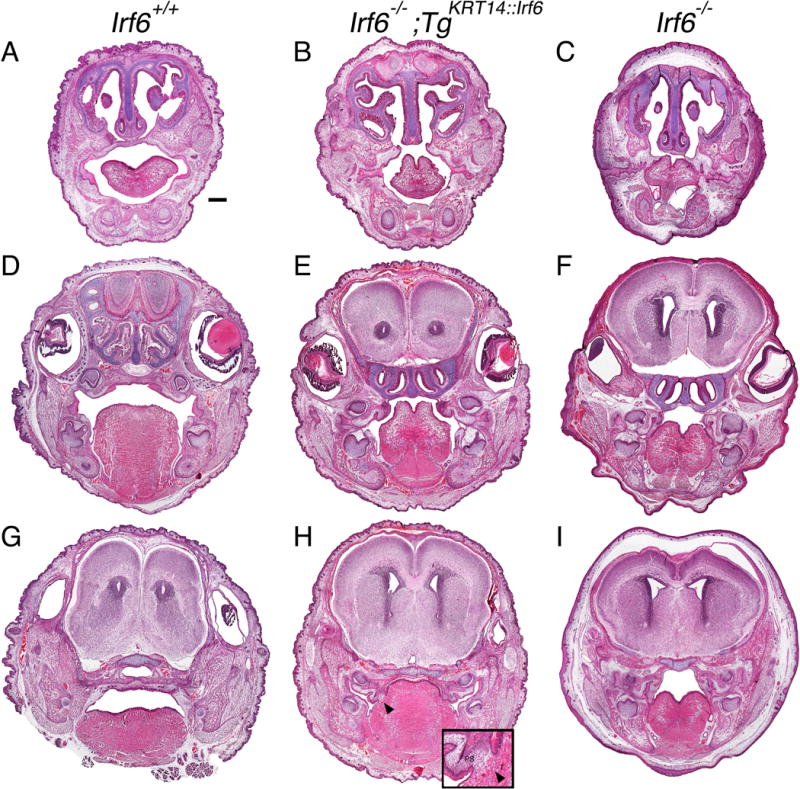
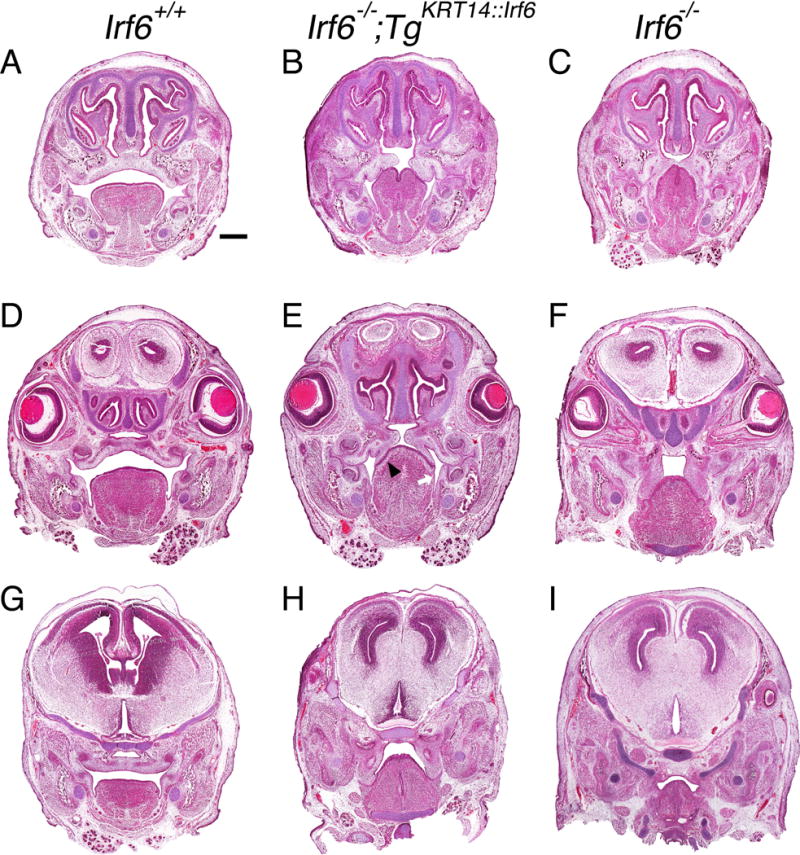
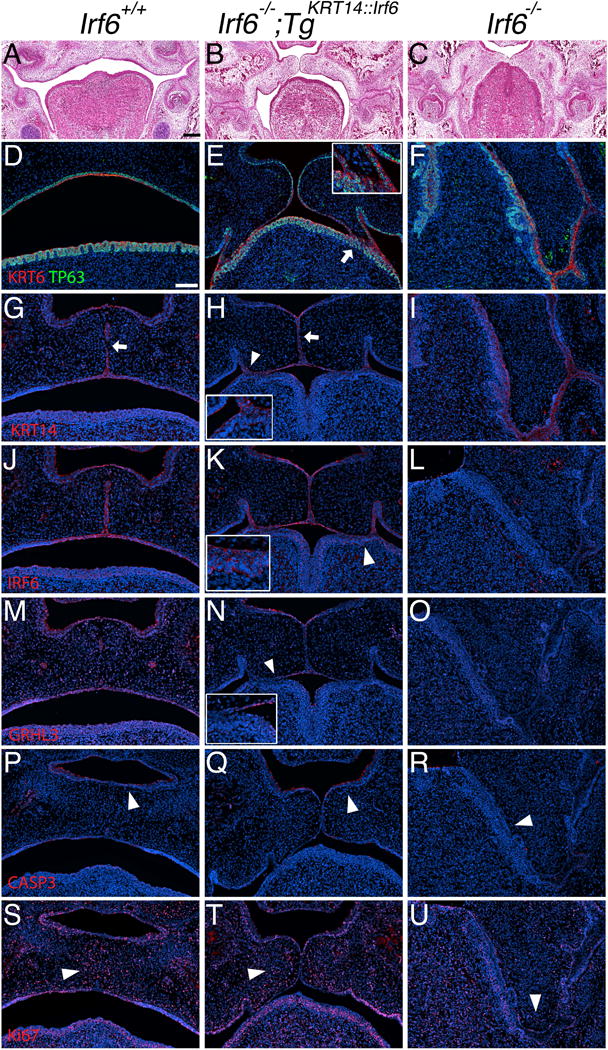
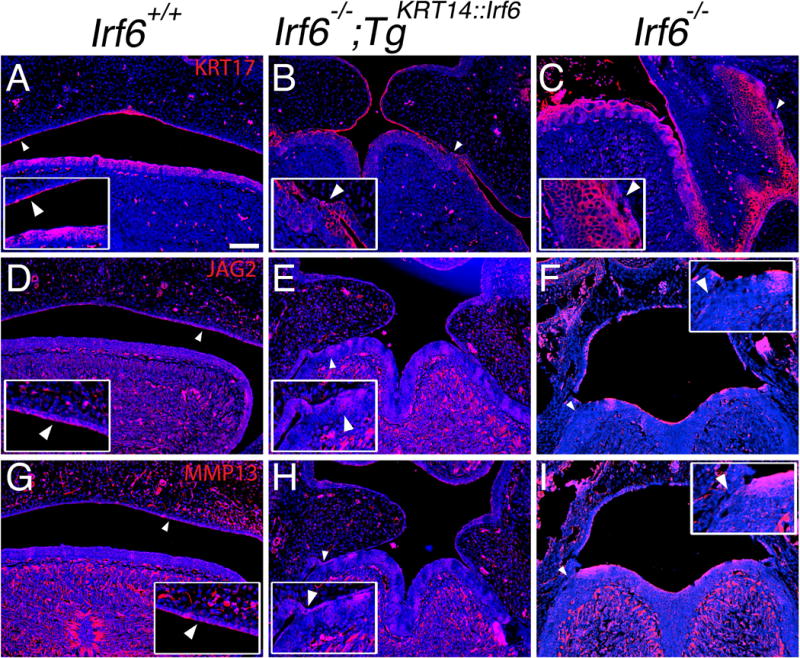
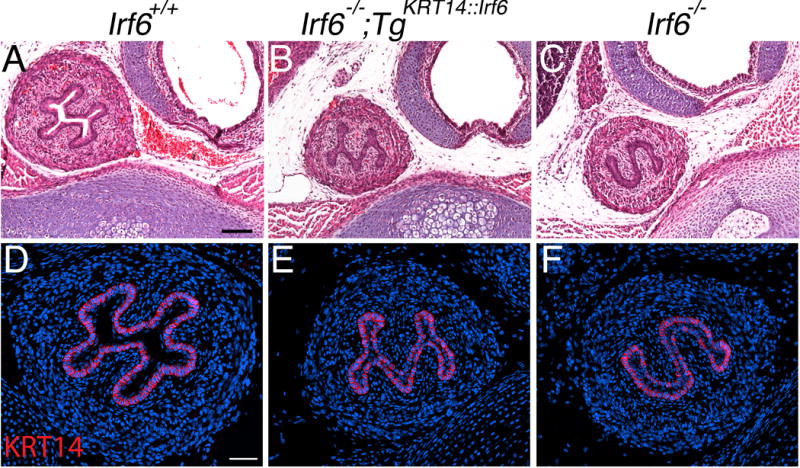
Results: Here, we show partial genetic rescue of Irf6 knockout embryos using the KRT14 promoter to drive Irf6 expression in the basal epithelium. In contrast to Irf6 knockout embryos, rescue embryos survive the immediate perinatal period. Macroscopic examination reveals rescue of skin adhesions between the axial and appendicular skeleton. Unexpectedly, KRT14-driven Irf6 expression does not completely rescue orofacial clefting and adhesions between the palate and tongue, suggesting the importance of cell-autonomous IRF6 expression in periderm. Like knockout embryos, Irf6 rescue embryos also have persistent esophageal adhesions, which likely contribute to postnatal demise.
Conclusions: Together, these data suggest that targeted expression of IRF6 can significantly reduce disease severity, but that a minimum level of Irf6 in both periderm and basal epithelial cells is necessary for orofacial development. Therefore, homologous human and mouse phenotypes are observed for IRF6, IKKA, and RIPK4. In this work, we show that altering the expression level of IRF6 dramatically modified this phenotype in utero. Developmental Dynamics 246:670-681, 2017. © 2017 Wiley Periodicals, Inc.
Keywords: GRHL3; IRF6; Van der Woude syndrome; cleft lip and palate; epidermis; epithelium; genetic rescue; limb defects; mouse models; oral periderm; popliteal pterygium syndrome.
© 2017 Wiley Periodicals, Inc.
Figures
References
-
- BOTTI E, SPALLONE G, MORETTI F, MARINARI B, PINETTI V, GALANTI S, DE MEO PD, DE NICOLA F, GANCI F, CASTRIGNANO T, PESOLE G, CHIMENTI S, GUERRINI L, FANCIULLI M, BLANDINO G, KARIN M, COSTANZO A. Developmental factor IRF6 exhibits tumor suppressor activity in squamous cell carcinomas. Proc Natl Acad Sci U S A. 2011;108:13710–5. - PMC - PubMed
-
- DE LA GARZA G, SCHLEIFFARTH JR, DUNNWALD M, MANKAD A, WEIRATHER JL, BONDE G, BUTCHER S, MANSOUR TA, KOUSA YA, FUKAZAWA CF, HOUSTON DW, MANAK JR, SCHUTTE BC, WAGNER DS, CORNELL RA. Interferon regulatory factor 6 promotes differentiation of the periderm by activating expression of grainyhead-like 3. J Invest Dermatol. 2013;133:68–77. - PMC - PubMed
-
- FAKHOURI WD, RAHIMOV F, ATTANASIO C, KOUWENHOVEN EN, FERREIRA DE LIMA RL, FELIX TM, NITSCHKE L, HUVER D, BARRONS J, KOUSA YA, LESLIE E, PENNACCHIO LA, VAN BOKHOVEN H, VISEL A, ZHOU H, MURRAY JC, SCHUTTE BC. An etiologic regulatory mutation in IRF6 with loss- and gain-of-function effects. Hum Mol Genet. 2014;23:2711–20. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
