

Post-translational modification of ribosomally synthesized peptides by a radical SAM epimerase in Bacillus subtilis
- PMID: 28644475
- PMCID: PMC6485343
- DOI: 10.1038/nchem.2714
Post-translational modification of ribosomally synthesized peptides by a radical SAM epimerase in Bacillus subtilis
Abstract
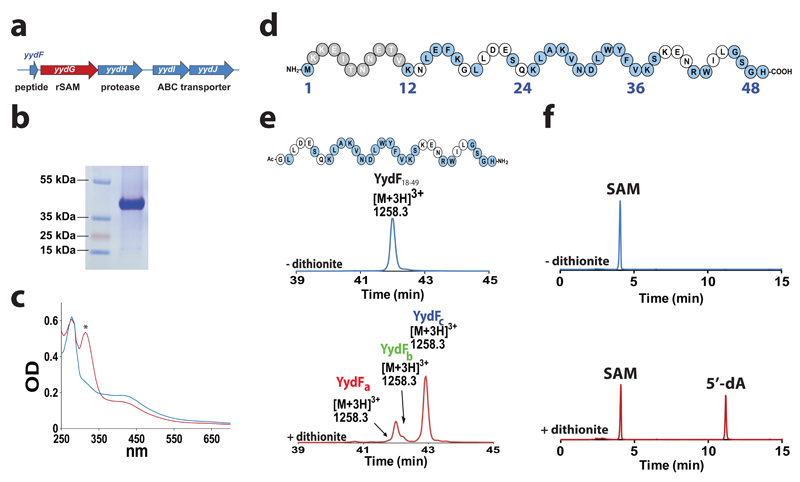
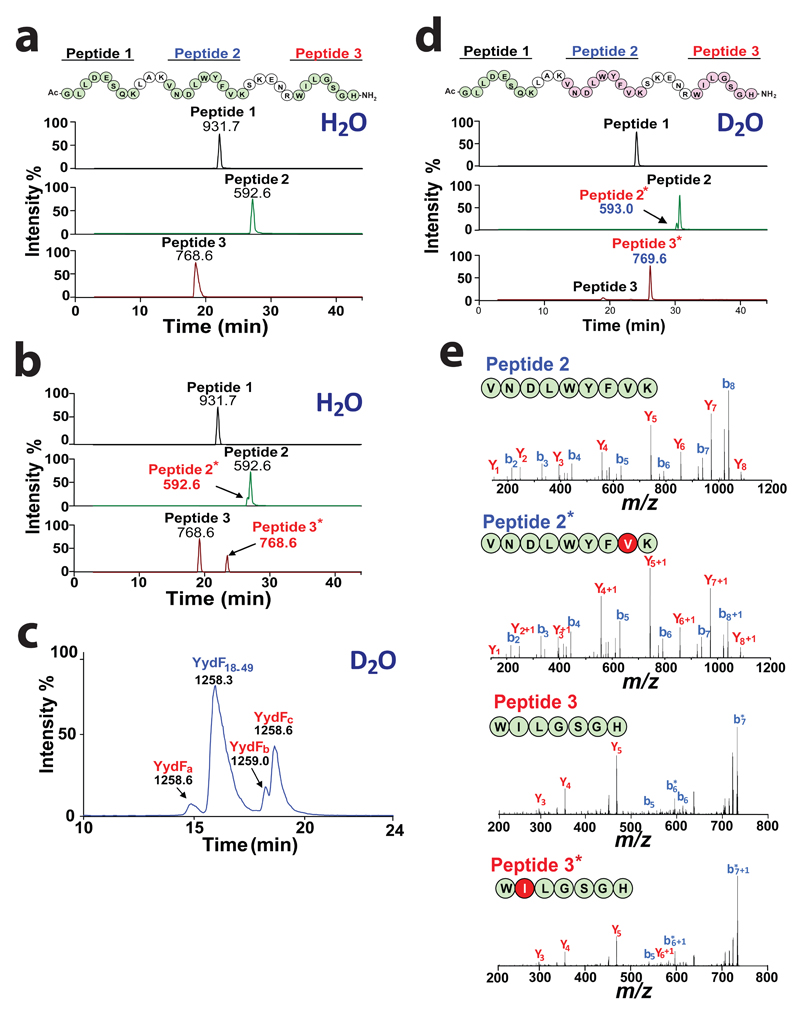
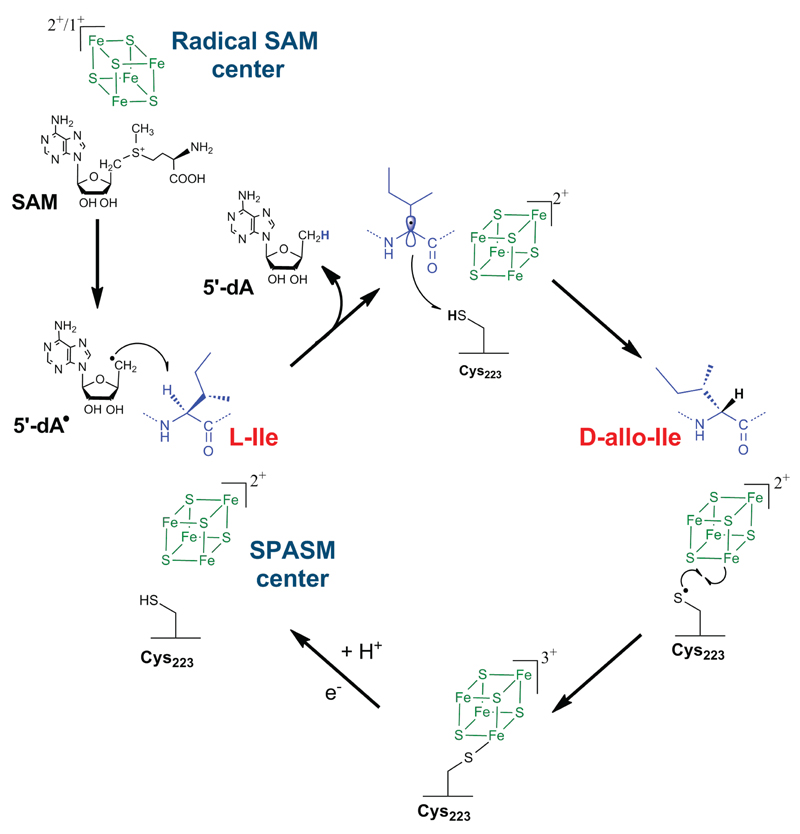
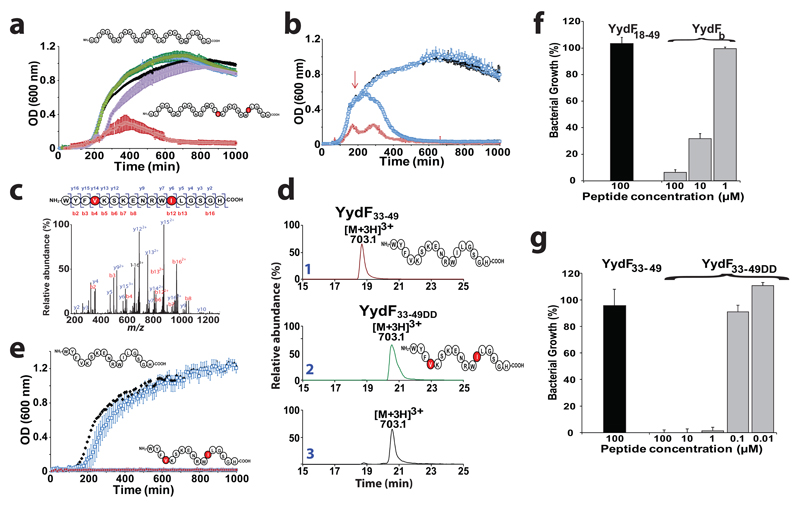
Ribosomally synthesized peptides are built out of L-amino acids, whereas D-amino acids are generally the hallmark of non-ribosomal synthetic processes. Here we show that the model bacterium Bacillus subtilis is able to produce a novel type of ribosomally synthesized and post-translationally modified peptide that contains D-amino acids, and which we propose to call epipeptides. We demonstrate that a two [4Fe-4S]-cluster radical S-adenosyl-L-methionine (SAM) enzyme converts L-amino acids into their D-counterparts by catalysing Cα-hydrogen-atom abstraction and using a critical cysteine residue as the hydrogen-atom donor. Unexpectedly, these D-amino acid residues proved to be essential for the activity of a peptide that induces the expression of LiaRS, a major component of the bacterial cell envelope stress-response system. Present in B. subtilis and in several members of the human microbiome, these epipeptides and radical SAM epimerases broaden the landscape of peptidyl structures accessible to living organisms.
Conflict of interest statement
The authors declare no competing financial interests
Figures


References
-
- Jordan S, Junker A, Helmann JD, Mascher T. Regulation of LiaRS-dependent gene expression in bacillus subtilis: identification of inhibitor proteins, regulator binding sites, and target genes of a conserved cell envelope stress-sensing two-component system. J Bacteriol. 2006;188:5153–66. - PMC - PubMed
-
- Jordan S, Hutchings MI, Mascher T. Cell envelope stress response in Gram-positive bacteria. FEMS Microbiol Rev. 2008;32:107–46. - PubMed
-
- Mascher T, Margulis NG, Wang T, Ye RW, Helmann JD. Cell wall stress responses in Bacillus subtilis: the regulatory network of the bacitracin stimulon. Mol Microbiol. 2003;50:1591–604. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
